2026年5月21日
理化学研究所
バイオ産業情報化コンソーシアム
東京大学
ダイナミックな相互作用が駆動する相分離機構
-溶液NMR法で明らかにした天然変性領域の役割-
理化学研究所(理研)生命医科学研究センター 生体分子動的構造研究チーム(研究当時)の嶋 田一夫 チームディレクター(研究当時)(バイオ産業情報化コンソーシアム(JBIC)特別顧問(研究当時)、現 理研生命医科学研究センター NMR共用促進チーム 客員主管研究員)、東京大学 大学院薬学系研究科 生命物理化学教室の竹内 恒 教授、同研究科 附属ワンストップ創薬共用ファシリティセンターの岡部 弘 基特任准教授らの共同研究グループは、RNAヘリカーゼ[1]の一種DEAD-Box RNA helicase 3 X-linked(DDX3X)[1]タンパク質が、細胞内のストレス顆粒[2]の形成を駆動する仕組みの一端を明らかにしました。
この研究成果は、ストレス顆粒のようにこれまでに創薬標的とは考えられていなかった非膜細胞小器官(オルガネラ)に対して作用するような、全く新しい機構を持つ創薬の技術開発に貢献すると期待されます。
DDX3Xは、mRNAの高次構造[3]をほどくRNAヘリカーゼ活性を持ち、細胞内のストレス顆粒の形成を制御する中心的な因子であることが知られています。しかし、DDX3Xのどのような分子間相互作用がストレス顆粒形成を制御しているのか、またDDX3XによるRNAの認識がストレス顆粒の機能にいかに寄与しているのか、詳細な分子機構は十分に明らかになっていませんでした。
今回、共同研究グループは、溶液核磁気共鳴分光法(溶液NMR法)[4]を用いて、DDX3Xの天然変性領域(IDR)[5]による、RNA中のグアニン4重鎖(GQ)構造[6]の選択的な認識が、ストレス顆粒形成、および顆粒へのDDX3XとRNAの局在を決定する一因であることを明らかにしました。
本研究は、科学雑誌『Journal of the American Chemical Society』オンライン版(5月20日付)に掲載されました。
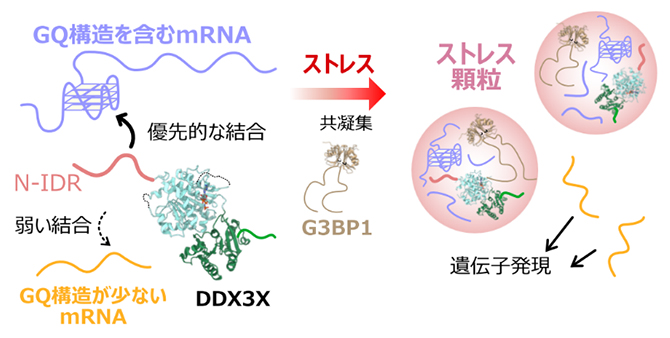
DDX3XがmRNAを選択的に認識し、翻訳制御を行う分子機構の模式図
背景
生物は細胞の内部に区画化された細胞小器官(オルガネラ)を形成し、生体分子の局在や輸送を通じて化学反応を制御することにより、生命活動を維持します。このようなオルガネラの形成は、古典的には脂質二重膜構造によって隔てられたオルガネラにより担われると考えられてきましたが、近年では、膜構造を持たない非膜オルガネラが多様な生命現象の制御に関与することが明らかになってきています。非膜オルガネラの多くは、タンパク質やRNA分子が水と油のように液-液相分離(LLPS)[7]を生じ、タンパク質とRNAが高度に濃縮された凝集相(液滴)を形成することにより細胞内の区画化を達成していると考えられています。このような液-液相分離を介した区画化は、形成や解離が迅速かつ可逆的に生じることから、生命反応の精緻な制御を行う上で重要な性質です。
非膜オルガネラの一例として、熱ショックや化学物質などのストレス依存的に細胞質において一過的に形成されるストレス顆粒が挙げられます。ストレス顆粒はタンパク質をコードするmRNAや、翻訳に関わるRNA結合タンパク質が、ストレス刺激依存的にLLPSを生じることによって形成される非膜オルガネラの一種です。ストレス顆粒は、ストレス環境下で翻訳を一時的に停止することで細胞生存応答の役割を担っており、その異常形成ががんや神経変性疾患にも関与することが知られています。従って、ストレス顆粒形成にまつわる分子機構を理解することは重要な課題でありながら、その形成がどのような生体分子間の相互作用により駆動されているのか、そしてどのような特徴を持つRNAが顆粒形成の促進、および顆粒への局在に重要であるのかは、十分に明らかになっていませんでした。
DEAD-Box RNA helicase 3 X-linked(DDX3X)は、RNAヘリカーゼの中でも最も大きなタンパク質ファミリーであるDEAD-Box RNAヘリカーゼの一種であり、mRNAの高次構造をほどくことで特定の遺伝子の翻訳促進をつかさどるタンパク質です。DDX3Xはストレス顆粒に局在するのみならず、その顆粒形成の鍵となる制御因子であることが知られています。共同研究グループはこれまでにDDX3XによるmRNAの認識機構を明らかにする研究に取り組み、DDX3Xの配列中で特定の構造を持たない天然変性領域(IDR)が、特徴的なRNAの高次構造の一種であるグアニン4重鎖(GQ)を持つRNA(GQ RNA)を認識することが、選択性を決定する要因であることを突き止めました注)。そこで共同研究グループは、これらのIDRとGQ RNAの相互作用が、ストレス顆粒形成においても重要な役割を果たすのではないかと着想し、その機能的な意義を詳細に調べることにしました。
- 注1)2025年8月28日プレスリリース「RNAを見分けてほどく、ヘリカーゼの分子機構」
研究手法と成果
今回、共同研究グループは、ヒト由来DDX3Xタンパク質を対象とし、RNAの結合に重要であることが既に分かっている、N末端[8]の天然変性領域(N-IDR)に着目し、非膜オルガネラ形成を駆動するLLPS活性を調べることにしました(図1A)。蛍光標識を施したN-IDRの蛍光顕微鏡を用いたイメージング解析から、N-IDRは、分子混雑環境(高濃度タンパク質の状態)を模倣するクラウダー分子存在下でLLPSを生じ(図1B)、形成されたタンパク質凝集相の液滴は、非膜オルガネラの特徴である高い流動性を示すことを光退色後蛍光回復法(FRAP法)[9]により明らかにしました(図1C)。
N-IDRのLLPS活性をつかさどる分子間相互作用を明らかにするために、溶液NMR法を用いて分子間相互作用に関与する部位を調べました。その結果、N-IDRの配列の中でも特に分子中央からC末端[8]にかけて、アルギニン残基や芳香族アミノ酸を多く含む領域(Arginine-rich region:RR領域)の相互作用が重要であること、そしてこれらの相互作用は弱く、かつ多価で働くダイナミックな相互作用様式であることを明らかにしました(図1D)。これらの残基の重要性は、RR領域のアルギニンをリジンに置換した変異体や、鍵となる芳香族アミノ酸をアラニンに置換したN-IDR変異体において、LLPS活性が低下していることからも確かめられました(図1E、F)。
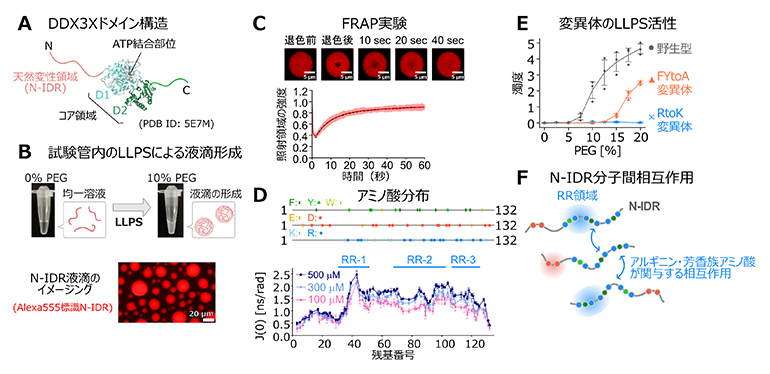
図1 DDX3Xタンパク質の構造とN-IDRのLLPS活性、ならびに分子間相互作用の評価
- A.DDX3Xドメイン構造。コア領域の立体構造はデータベース上で公開されているもの(PDB ID: 5E7M)を参照し、N、C末端の天然変性領域、コア領域(D1、D2)を色分けして示した。
- B.試験管内のN-IDR溶液に分子混雑環境を模倣するためにクラウダー分子(PEG)を添加したときの溶液の変化。LLPSに伴い液滴が形成されたことに起因する溶液の白濁が観測された。蛍光色素(Alexa555)で標識したN-IDRの液滴の蛍光顕微鏡による観察から、液滴構造を確認した。
- C.蛍光標識N-IDRのFRAP実験の結果。(上)レーザーによる退色後、退色部位に周囲から分子が流入する様子を捉えた共焦点画像、(下)退色部位のシグナル強度の時間経過の変化をプロットしたもの。退色部位の蛍光が時間が経つと元の強度に回復したことから、液滴は流動性を有していることが確認された。
- D.(上)N末端の天然変性領域(N-IDR)を形成する132個のアミノ酸残基の配列上にある芳香族アミノ酸(F: フェニルアラニン、Y:チロシン、W:トリプトファン)、負電荷を帯びたアミノ酸(E:グルタミン酸、D:アスパラギン酸)、および正電荷を帯びたアミノ酸(K:リジン、R:アルギニン)の分布を示した。(下)N-IDRの濃度を変化させたときの、分子間相互作用に伴う運動性の低下の度合いを反映するスペクトル密度成分(J(0 ))(分子運動性を評価する上で、特定の周波数に対応する成分がどの程度の強さで分布しているかを示す関数のうちゼロ周波数の成分、ラジアン分のナノ秒(10億分の1秒)[ns/rad] の単位で表され、値が大きいほど運動性が低下していることを示唆する)をプロットしたNMR実験の結果。高濃度条件において顕著な運動性の低下が観測された領域から、主要な相互作用領域を同定した。これらの相互作用領域は、配列上でアルギニン残基が集積したクラスター領域と一致することが分かった。μM:マイクロモーラー(100万分の1モーラー(M))
- E.N-IDR変異体のLLPSアッセイの結果。クラウダー分子(PEG)の濃度を増大させたときの、溶液の濁度の増大から液滴形成を評価した。RR領域のフェニルアラニン(F)、チロシン(Y)を変異させたFYtoA変異体、およびアルギニン(R)をリジン(K)に変異させたRtoK変異体では顕著にLLPS活性が低下していることが分かった。
- F.NMR実験、およびLLPSアッセイから明らかとなったN-IDR分子間相互作用を模式的に表した図。
次に、N-IDRによるGQ RNAの認識がLLPS活性にどのように影響するかを詳細に調べました。蛍光標識を施したN-IDRとGQ RNAを混合したところ、クラウダー分子非存在下においてもタンパク質凝集相の液滴が生じること、液滴内部にはN-IDRとGQが共局在していることが分かりました(図2A上・中)。興味深いことに、GQ構造を持たない変異配列から成るRNAを用いて同様の実験を行ったところ、LLPSが観測されませんでした(図2A下)。このことは、GQ RNAを用いた際に観測されたLLPS活性は、N-IDRによるGQ RNAの構造特異的認識により担われていることを強く支持する結果です。さらに溶液NMR法を用いて、N-IDR分子間の相互作用を、GQ RNAの存在下、非存在下で比較しました(図2B)。解析の結果、RR領域で生じる分子間相互作用がGQ RNA存在下で顕著に増大していたことから、GQ RNAはRR領域で生じる分子間相互作用を促進することによりLLPSを効率的に誘起していることが示唆されました(図2C)。
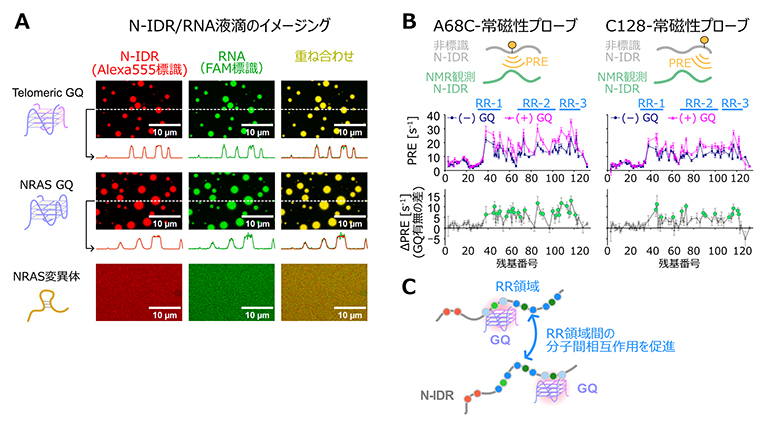
図2 N-IDRのGQ RNA依存的なLLPSと分子間相互作用の変化の解析
- A.蛍光色素で標識したN-IDR(Alexa555、赤色)、およびRNA(FAM、緑色)から成る液滴の共焦点顕微鏡による観察の結果。(上)Telomeric(テロメア) GQ RNA、(中)NRAS(がん遺伝子であるRASファミリーの遺伝子の一種)GQ、および(下)NRAS変異体と異なるRNA配列を用いて比較した結果を示す。
- B.N-IDR分子間相互作用を検出するための分子間PRE(paramagnetic relaxation enhancement)実験の結果。PRE実験とは、部位特異的に導入した常磁性プローブ(常磁性を示す不対電子対を持つ化合物)からの近接効果を、NMR観測核のシグナルの緩和(磁化の減衰)が促進する現象を利用して定量する。分子間PRE実験では、常磁性プローブを導入した非標識のN-IDR分子から、安定同位体標識することでNMR観測を可能としたN-IDR分子に生じる緩和促進効果を解析する。(上)残基番号68(左)、および128(右)に常磁性プローブを導入した時の分子間PRE実験の模式図。(中)GQ RNA存在下(マゼンタ)、非存在下(紺)におけるPRE、およびその差分のプロット。顕著な(> 5毎秒)PREが観測された残基は緑色の球でハイライトした。
- C.分子間PRE実験から明らかとなったN-IDR分子間相互作用へのGQ RNAの寄与を模式的に表した図。
このようなN-IDRとGQ RNAから成るLLPS現象が生理的なストレス顆粒形成を反映したものであるかを調べるために、ストレス顆粒のコア因子であるG3BP1タンパク質[10]と共凝集し、同じ液滴内に局在する共LLPS活性を調べました。蛍光標識を施したN-IDRとGQ RNA、およびG3BP1を混合したところ、ストレス顆粒を模したタンパク質凝集相の液滴が形成され、3者全てが液滴内部に共局在していることが確認されました(図3A上・中)。同様のLLPS現象は、GQを形成しない変異体配列のRNAを用いた際には観測されなかったことから(図3A下)、DDX3XのN-IDRとGQ RNAの相互作用はストレス顆粒へのRNAの局在を決定する一因であることが示唆されました。以上の結果を統合し、DDX3XのN-IDRによるGQ RNAの構造特異的な認識は、ストレス顆粒内に分配するRNAの選択性を決める一因であると提唱しました(図3B)。
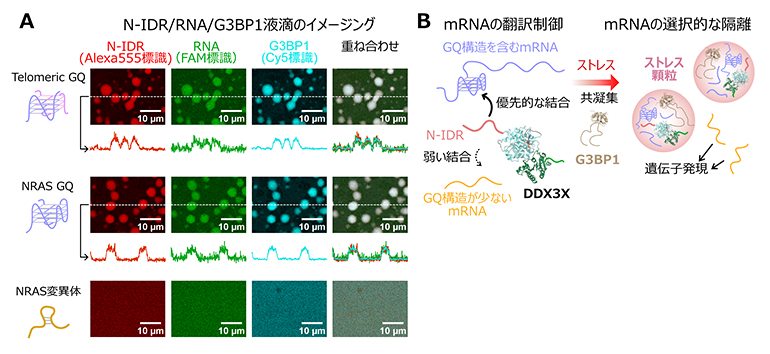
図3 N-IDRとRNA、G3BP1から成るLLPSの解析とストレス顆粒形成の翻訳制御機構
- A.蛍光色素で標識したN-IDR(Alexa555、赤色)、RNA(FAM、緑色)、およびG3BP1(Cy5、シアン)から成る液滴の共焦点顕微鏡による観察の結果。(上)Telomeric GQ RNA、(中)NRAS GQ、および(下)NRAS変異体と異なるRNA配列を用いて比較した結果を示す。
- B.本研究成果で得られたDDX3Xの分子機構の模式図。DDX3XのN-IDRがmRNAのGQ構造を選択的に認識し、ストレス環境下でG3BP1と共凝集することでストレス顆粒を形成する。GQ構造を持たないmRNAは顆粒内から排斥されることで、ストレス環境下でも遺伝子発現が維持される。
今後の期待
本研究では、RNAヘリカーゼDDX3XのN-IDRに着目し、N-IDRによるGQ RNAの構造特異的認識がLLPS活性、および生理的なストレス顆粒へのRNAの局在を決定する一因であることを明らかにしました。特にこれらのLLPS活性は、弱く多価的であることを特徴とするダイナミックな分子間相互作用により担われていることから、これらの相互作用を検出することが可能な溶液NMR法の重要性を強く支持するものです。
最近の研究では、ヒトのプロテオーム(全タンパク質種)の中でも~20%近くもの割合が非膜オルガネラ形成に関与していると提唱されており、非膜オルガネラの形成にまつわる分子機構を理解することは、複雑かつ精緻な生命現象を理解する上でますます重要になると注目を集めています。特に、溶液NMR法から得られる分子・原子レベルでの知見は、ストレス顆粒のようにこれまでに創薬標的とは考えられていなかった非膜オルガネラに対して作用するような、全く新しい機構を持つ創薬技術の開発に貢献すると期待されます。
補足説明
- 1.RNAヘリカーゼ、DEAD-Box RNA helicase 3 X-linked(DDX3X)
RNAヘリカーゼは、RNAのヘリックス(らせん)構造などの高次構造を解消する機能を持つ酵素。DEAD-Box RNAヘリカーゼは、活性部位にアスパラギン酸-グルタミン酸-アラニン-アスパラギン酸(D-E-A-D)から成る保存された配列モチーフを持つことを特徴とする、RNAの高次構造([3]参照)をアデノシン三リン酸(ATP)依存的にほどくRNAヘリカーゼのファミリー。ヒトにおいて最も大きなファミリーを構成する。DEAD-Box RNA helicase 3 X-linked(DDX3X)はX染色体上にその遺伝子が存在するDEAD-Box RNAヘリカーゼの一種である。 - 2.ストレス顆粒
熱ショックや酸化ストレスなどの刺激を受けた際、細胞質内においてmRNAとタンパク質が共凝集することで、一過的に形成される非膜オルガネラ。mRNAや翻訳開始因子、RNA結合タンパク質を取り込み隔離することで、ストレス環境下における翻訳プロファイルを制御し、細胞の生存応答に寄与する。ストレスが除去されると速やかに解消される可逆的な性質を持つが、慢性的ストレスなどの要因により、ストレス顆粒の過剰形成や固体状への成熟が進むと、神経変性疾患に関与する毒性凝集体の蓄積の場となる。 - 3.高次構造
アデノシン(A)、シチジン(C)、グアノシン(G)、ウリジン(U)の4種類のリボヌクレオシドから構成されるRNAが、分子間もしくは分子内で相互作用することで形成される立体構造をRNAの高次構造と呼ぶ。一部の遺伝子産物のmRNAの5'末端の非翻訳領域は2本鎖やループ、グアニン4重鎖(GQ)構造([6]参照)などさまざまな高次構造を形成することが知られており、これらの高次構造形成は翻訳マシナリー(装置)の結合や機能を阻害することで、翻訳を抑制的に制御する。RNAヘリカーゼは、これらの高次構造をほどくことによって、特定の遺伝子の翻訳を促進する。 - 4.溶液核磁気共鳴分光法(溶液NMR法)
溶液中の化合物や生体分子などを対象に、強い磁場中に置くことで生じる原子核の共鳴現象を観測することで、その化学構造や運動性といった情報を原子レベルで解析できる分光法。磁場の中に置いた試料に電磁波を照射し、そこから放出される電磁波をNMRシグナルとして検出する。NMRはNuclear Magnetic Resonanceの略。 - 5.天然変性領域(IDR)
タンパク質の中でもαヘリックスやβシートなどのような固有の高次構造を形成しない領域。IDRはヒトのプロテオームの30%以上を占めるという試算もあることから、多くのタンパク質がIDRを有していると考えられているものの、IDRが機能に果たす役割は明らかになっていないケースも多い。IDRはIntrinsically Disordered Regionの略。 - 6.グアニン4重鎖(GQ)構造
DNAやRNAにおいて、四つのグアニン塩基が同じ平面上で、ワトソン・クリック型塩基対(アデニンとチミンもしくはウラシル、グアニンとシトシンとの間で水素結合が形成される典型的な塩基対)とは異なるフーグスティーン型塩基対(塩基が回転することで、グアニン間で形成される非典型的な塩基対)を形成し、重なり合うことで形成される特徴的な高次構造。グアニン塩基の中央には、カリウムイオンなどの1価の陽イオンが配位することで安定化に寄与する。GQはG-quadruplexの略。 - 7.液-液相分離(LLPS)
高濃度条件や分子混雑環境においてタンパク質や核酸が凝集し、周囲から区画化された「凝集相(液滴)」と、その外部の「希薄相」の2相に分離する現象。水と油が分離する相分離現象に近く、特定の分子を局所的に濃縮することで、非膜オルガネラ形成の基盤となる物理現象であると考えられている。形成された凝集相は、内部で高い拡散性を保ちつつ、希薄相と分子をやり取りする流動的な性質を持つ。LLPSはLiquid-Liquid Phase Separationの略。 - 8.N末端、C末端
タンパク質はアミノ酸残基が直鎖状に連結したポリペプチドで構成されることから、その始まりと終わりがある。タンパク質の始まりとなる1番目の残基(またはその主鎖アミノ基)がN末端、終わりとなる最後の残基(またはその主鎖カルボキシル基)がC末端と呼ばれる。 - 9.光退色後蛍光回復法(FRAP法)
試験管内や細胞内にて形成される蛍光標識分子を含む非膜オルガネラや脂質二重膜構造などの構造体において、分子の流動性を評価する手法。強いレーザー照射により一部の蛍光分子を局所的に退色させた後、周囲から分子が拡散することで蛍光が回復する過程を測定する。回復速度を解析することで、内部における分子の拡散係数を定量的に評価することが可能である。FRAPはFluorescence Recovery After Photobleachingの略。 - 10.G3BP1タンパク質
ストレス顆粒の形成において中心的な役割を果たすRNA結合タンパク質の一種。G3BP1を欠損させると、ストレス顆粒の形成が著しく阻害されることから、ストレス顆粒形成の足場として機能する分子であるとされている。分子内にRNA結合ドメインや天然変性領域を持ち、これらを介してmRNAや他のタンパク質と多価的な相互作用を形成し、LLPSを駆動するスイッチ分子として機能する。G3BP1はRas GTPase-activating protein SH3 domain binding protein 1の略。
共同研究グループ
理化学研究所 生命医科学研究センター
生体分子動的構造研究チーム(研究当時)
チームディレクター(研究当時)嶋田 一夫(シマダ・イチオ)
(現 バイオ産業情報化コンソーシアム(JBIC)理事、理研名誉研究員、生命医科学研究センター NMR共用促進チーム 客員主管研究員、広島大学 副学長)
客員研究員(研究当時)外山 侑樹(トヤマ・ユウキ)
(現 東京大学 大学院薬学系研究科 特任助教、理研 生命医科学研究センター 細胞内構造ダイナミクス研究チーム 客員研究員)
東京大学 大学院薬学系研究科
生命物理化学教室
教授 竹内 恒(タケウチ・コウ)
大学院生 稲上 慎一郎(イナカミ・シンイチロウ)
附属ワンストップ創薬共用ファシリティセンター
特任准教授 岡部 弘基(オカベ・コウキ)
特任助教 寳田 雅治(タカラダ・マサハル)
研究支援
本研究は、日本医療研究開発機構(AMED)「次世代治療・診断実現のための創薬基盤技術開発事業(RNA標的創薬技術開発)」による助成を受けて行われました。蛍光顕微鏡を用いた解析は、理研環境資源科学研究センターの質量分析・顕微鏡解析ユニットの支援の下で、共焦点顕微鏡を用いた解析は東京大学大学院薬学系研究科附属ワンストップ創薬共用ファシリティセンターの支援の下で実施されました。
原論文情報
- Yuki Toyama, Shinichiro Inakami, Masaharu Takarada, Kohki Okabe, Koh Takeuchi, Ichio Shimada, "Phase separation driven by dynamic interactions in the N-terminal intrinsically disordered region of the DEAD-box RNA helicase DDX3X", Journal of the American Chemical Society, 10.1021/jacs.5c22149
発表者
理化学研究所
生命医科学研究センター 生体分子動的構造研究チーム(研究当時)
チームディレクター(研究当時)嶋田 一夫(シマダ・イチオ)
(バイオ産業情報化コンソーシアム(JBIC)特別顧問(研究当時)、現 理研 生命医科学研究センター NMR共用促進チーム 客員主管研究員)
東京大学 大学院薬学系研究科
生命物理化学教室
教授 竹内 恒(タケウチ・コウ)
附属ワンストップ創薬共用ファシリティセンター
特任准教授 岡部 弘基(オカベ・コウキ)
報道担当
理化学研究所 広報部 報道担当
お問い合わせフォーム
バイオ産業情報化コンソーシアム 問い合わせ窓口
JBICお問い合わせフォーム
東京大学 大学院薬学系研究科 庶務チーム
Tel: 03-5841-4702
Email: shomu@mol.f.u-tokyo.ac.jp