理化学研究所(理研)開拓研究所 平野染色体ダイナミクス研究室(研究当時)の新冨 圭史 専任研究員(研究当時、現 生命医科学研究センター 転写制御構造生物学研究チーム 専任研究員)と平野 達也 主任研究員(研究当時、現 名誉研究員)らの研究チームは、独自に開発した染色体再構成系[1]を駆使し、主要な染色体構成タンパク質複合体であるコンデンシンⅠ[2]が機能のオンとオフを切り替える仕組みを明らかにしました。
本研究成果は、細胞増殖の基本原理の理解に貢献すると期待されます。
細胞分裂の前後で、遺伝情報を継承するために染色体は「構築」と「脱構築」を繰り返します。この重要かつダイナミックな構造変化を制御する仕組みは、細胞生物学における長年の疑問でした。
今回、研究チームは、最小限の精製タンパク質を用い、試験管内で染色体の構築と脱構築のプロセスを再現する実験手法を確立しました。その手法により、「2段階のリン酸化」によってもたらされるコンデンシンⅠの活性化が染色体構築に不可欠であることを見いだしました。一方、染色体脱構築の引き金を引くのは、コンデンシンⅠの脱リン酸化であることも証明しました。
本研究は、科学雑誌『PNAS Nexus』オンライン版(6月25日付)に掲載されました。
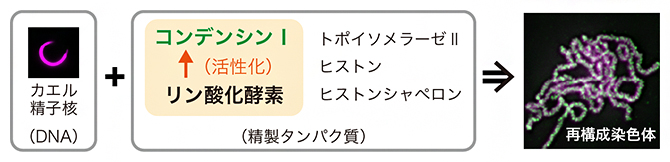
精製タンパク質を使った染色体再構成
背景
細胞分裂の直前の時期(分裂期)には、核内に広がって存在するクロマチン[3]から棒状の染色体が形成されます。そして分裂期が終了すると、染色体は再び広がったクロマチンへと戻ります。この染色体の「構築」と「脱構築」は、遺伝情報の正確な継承と適切な遺伝子発現に不可欠なプロセスです。
この現象の分子的理解を大きく前進させたのが、1997年にカエル卵抽出液[4]を用いた研究から発見されたコンデンシンⅠです。その後の研究によって、コンデンシンⅠが分裂期染色体の構築を担う中心的なタンパク質複合体であることが明らかになりました。さらに近年では、コンデンシンⅠがDNA上にループ構造をつくり出し、それらを秩序立って配置することで染色体を形成することが分かってきました。
一方で、コンデンシンⅠは細胞内に常に存在しているにもかかわらず、実際に機能するのは分裂期に限られています。つまり、分裂期の開始とともに活性化され、終了とともに不活性化されると考えられます。しかし、コンデンシンⅠがどのようにしてオンとオフを切り替え、染色体の構築と脱構築を制御しているのかは、その発見から約30年が経った現在でも十分に理解されていませんでした。研究チームは、限られた数の精製因子のみを用いて染色体を再構成する独自の手法を活用し、この長年の謎の解明に挑みました。
研究手法と成果
コンデンシンⅠの活性調節の仕組みを理解する手がかりは、新冨専任研究員らが2015年に開発した染色体再構成法注1)にありました。この方法では、わずか6種類の精製タンパク質とDNA(正確にはアフリカツメガエルの精子核)から、分裂期染色体を試験管内で再構成できます注2)。当時、染色体再構成系に用いられていたコンデンシンⅠは、分裂期で停止したアフリカツメガエル未受精卵から精製した「リン酸化済み」のものでした。また、それまでの研究成果注3、4)から、コンデンシンⅠのリン酸化が染色体構築に重要であり、リン酸化部位の多くはD2とHと呼ばれる二つのサブユニットの「天然変性領域[5]」に集中していると予想されていました。
今回の研究では、卵から精製したコンデンシンⅠの代わりに、人工的に作製した「組換え型」コンデンシンⅠを用い、その活性化過程を再現できるかを調べました。
まず、組換え型コンデンシンⅠは、ほとんどリン酸化されておらず、染色体構築を促進できないことを確認しました。一方、この組換え型コンデンシンⅠを細胞分裂の開始を制御する「リン酸化酵素Cdk1-サイクリンB複合体」で処理すると、染色体構築能を獲得し、卵から精製したコンデンシンⅠと同等の活性を示しました。さらに解析の結果、活性化されたコンデンシンⅠでは、複数のサブユニットの複数のアミノ酸がリン酸化されていることが分かりました。
次に、どの領域のリン酸化が活性化に重要なのかを明らかにするため、組換え型コンデンシンⅠのさまざまな変異体を作製し、それらの機能を染色体再構成系を用いて調べました。その結果、コンデンシンⅠの活性化には「2段階のリン酸化」が必要であることが明らかになりました。まず、分裂期に入ると、天然変性領域がリン酸化されます。すると、この領域が持つ自己抑制作用が解除され、コンデンシンⅠはDNAへ効率よく結合できるようになります。しかし、この段階だけでは染色体構築に十分ではありませんでした。さらに、天然変性領域以外(コア領域)のアミノ酸がリン酸化されることで、初めてコンデンシンⅠは最大限の活性を発揮し、染色体構築を効率よく進められることが分かりました。つまり、コンデンシンⅠは2種類のリン酸化スイッチを段階的に作動させることで、染色体構築活性を精密に制御していると考えられます。
一方、染色体の「脱構築」過程にも着目しました。分裂期終了時には、リン酸化酵素Cdk1-サイクリンBの活性が急速に低下し、代わって「脱リン酸化酵素PP2A-B55」が活性化されることが知られています。そこで、試験管内で染色体再構成が完了した反応液にPP2A-B55を加えたところ、コンデンシンⅠの天然変性領域が効率よく脱リン酸化されることが分かりました。さらに、コンデンシンⅠが染色体から離れ、それに伴って特徴的な棒状の染色体構造が崩れていく様子が観察されました。これは、染色体の脱構築過程を試験管内で初めて再現した成果です。
さらに、カエル卵抽出液を用いた解析によって、この段階的な活性化と不活性化の仕組みが細胞内でも実際に機能していることを確認しました。
これらの結果は、分裂期を通じた染色体のダイナミックな振る舞いが、リン酸化スイッチが連携して働く精巧な制御ネットワークによって実現されていること、そしてその中核がCdk1-サイクリンBとPP2A-B55という一組の酵素によって担われていることを示しています。
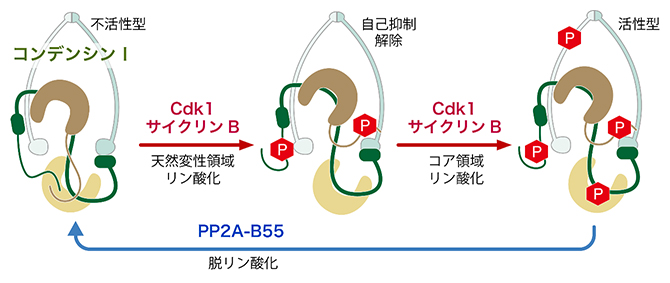
図1 リン酸化によるコンデンシンⅠの活性化と不活性化のモデル
活性化前(左)の第一段階から、コンデンシンⅠの天然変性領域がリン酸化され、自己抑制が解除される(中央)。第二段階として、コア領域がさらにリン酸化され、染色体構築を効率的に進められる活性型になる(右)。不活性化は脱リン酸化によってもたらされる。P:リン酸。
- 注1)Shintomi, K., Takahashi, T. S., and Hirano, T. "Reconstitution of mitotic chromatids with a minimum set of purified factors." Nat. Cell Biol. 17, 1014-1023 (2015).
- 注2)2021年5月18日プレスリリース「試験管内で染色体を作る」
- 注3)Kimura, K. et al. "Phosphorylation and activation of 13S condensin by Cdc2 in vitro." Science, 282, 487-90 (1998).
- 注4)Tane, S. et al., "Cell cycle-specific loading of condensin I is regulated by the N-terminal tail of its kleisin subunit." eLife, 11, e84694 (2022).
今後の期待
今回の研究によって、細胞分裂に伴う染色体の構築と脱構築を制御する基本原理の一端が明らかになりました。一方で、コンデンシンⅠがどのようにして染色体を形成するのか、その分子レベルでの動作原理には未解明な部分が多く残されています。近年の研究により、コンデンシンⅠは巨大なタンパク質複合体でありながら、柔軟に構造を変化させることが分かってきました。今後は、リン酸化と脱リン酸化によってコンデンシンⅠの構造がどのように変化し、その機能がどのように制御されるのかを理解することが重要な課題です。そのためには、多様な分野の研究者が協力し、最新の解析技術と染色体再構成系を組み合わせていくことが必要になると考えられます。
また、細胞現象を試験管内で再構成する研究手法は、複雑な生命現象を最小限の構成要素から理解するための強力なアプローチです。コンデンシンⅠのみならず、分裂期に働く多くのタンパク質は、リン酸化によってその機能が制御されていることが知られており、今回確立した材料や方法が、核膜崩壊と再構築、紡錘体(ぼうすいたい)形成、細胞質分裂などの、さまざまな分裂期現象の解析にも応用できる可能性があります。近年、多種多様な生命現象を試験管内で再現し、その仕組みを理解する研究手法は大きく発展しています。細胞のダイナミックな振る舞いを「つくって理解する」時代は、着実に近づいているようです。
補足説明
- 1.染色体再構成系
分裂期の細胞内で起こる染色体構築過程を、現在考え得る最小の要素を用いて試験管内で再現する実験系。2015年に、新冨専任研究員と平野主任研究員が報告した方法では、6種類の精製タンパク質(リン酸化型コンデンシンⅠ、トポイソメラーゼⅡ、コアヒストン、3種類のヒストンシャペロン)、アフリカツメガエルの精子核、ATP(アデノシン三リン酸)を水溶液中で混和し、22℃でインキュベーションする。 - 2.コンデンシンⅠ
染色体構築に中心的な役割を果たすタンパク質複合体。5種類のサブユニットから構成される。カエル卵抽出液([4]参照)でつくった染色体に含まれる主要なタンパク質成分として、1997年に同定された。その後、多くの真核生物で、サブユニットの一部が異なる複合体コンデンシンⅡの存在も明らかにされている。 - 3.クロマチン
真核生物のゲノムDNAは、さまざまなタンパク質と相互作用したクロマチンと呼ばれる構造として細胞内に存在している。その単位構造は、コアヒストン8量体にDNAが巻きついた「ヌクレオソーム」と呼ばれるものである。 - 4.カエル卵抽出液
カエル(アフリカツメガエル)の卵を遠心破砕して得られる抽出液。実質的には、希釈されずに外部に取り出された卵の可溶性成分であり、受精や細胞分裂周期の進行に伴う細胞内の生化学反応を試験管内で再現できる。1983年に、トロント大学(カナダ)の増井禎夫博士(1931~2024年)によって開発され、その後の細胞分裂周期研究の飛躍的発展の一翼を担った。 - 5.天然変性領域
タンパク質の構造のうち、生理的条件下で決まった立体構造(ヘリックス構造やシート構造:二次構造と呼ばれる)をつくらない領域。タンパク質同士の相互作用や核酸(DNAやRNA)との相互作用に関与し、タンパク質の機能を調節する。
研究チーム
理化学研究所 開拓研究所 平野染色体ダイナミクス研究室(研究当時)
専任研究員(研究当時)新冨 圭史(シントミ・ケイシ)
(現 生命医科学研究センター 転写制御構造生物学研究チーム 専任研究員)
特別研究員(研究当時)田根 将志(タネ・ショウジ)
テクニカルスタッフⅠ(研究当時)正原 由紀(マサハラ・ユキ)
主任研究員(研究当時、現 名誉研究員)平野 達也(ヒラノ・タツヤ)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費助成事業基盤研究(B)「染色体の構築・脱構築メカニズムの解明:コンデンシンⅠのリン酸化制御を手がかりに(研究代表者:新冨圭史、JP25K02211)」、同基盤研究(S)「コンデンシンⅠとⅡの分子メカニズムの解明(研究代表者:平野達也、JP18H05276)」、文部科学省科学研究費助成事業学術変革領域研究(A)「分裂期染色体の礎となるバイオロジカルクラスターの解析と再構成(研究代表者:新冨圭史、JP25H02407)」「分裂期染色体のモダリティ(研究代表者:平野達也、JP20H05938)」による助成を受けて行われました。
原論文情報
- Keishi Shintomi, Shoji Tane, Yuki Masahara-Negishi, and Tatsuya Hirano, "Dual phosphoregulatory mechanisms of condensin I revealed by biochemical reconstitution", PNAS Nexus, 10.1093/pnasnexus/pgag218
発表者
理化学研究所
開拓研究所 平野染色体ダイナミクス研究室(研究当時)
専任研究員(研究当時)新冨 圭史(シントミ・ケイシ)
(現 生命医科学研究センター 転写制御構造生物学研究チーム 専任研究員)
主任研究員(研究当時、現 名誉研究員)平野 達也(ヒラノ・タツヤ)
報道担当
理化学研究所 広報部 報道担当
お問い合わせフォーム