理化学研究所(理研)開拓研究所 眞貝細胞記憶研究室の福田 渓 客員研究員、志村 知古 テクニカルスタッフⅠ、眞貝 洋一 主任研究員らの共同研究グループは、胎盤が正常につくられるために欠かせない新たな制御因子「RESF1」の働きを明らかにしました。
本研究成果は、胎盤形成を支えるゲノムの働きがどのように制御されているのかという理解を大きく前進させるものであり、胎盤形成異常や妊娠関連疾患の原因解明につながることが期待されます。
胎盤は、母親から胎児へ酸素や栄養を供給し胎児から母体へ老廃物を排出する重要な臓器であり、胎児が健やかに成長するために欠かせません。その正常な形成には、胎盤の細胞でゲノムが適切に機能することが必要ですが、その仕組みは十分には分かっていませんでした。
共同研究グループは、これまで生体内での役割がほとんど分かっていなかったRESF1が、胎盤の細胞でゲノムの働きを適切な状態に維持し、正常な胎盤形成に欠かせない役割を果たしていることを明らかにしました。さらに、RESF1は従来知られていた仕組みとは異なる経路で機能することを示し、胎盤細胞に特有の新しいゲノム制御機構を明らかにしました。
本研究は、科学雑誌『Genome Research』オンライン版(6月29日付)に掲載されました。
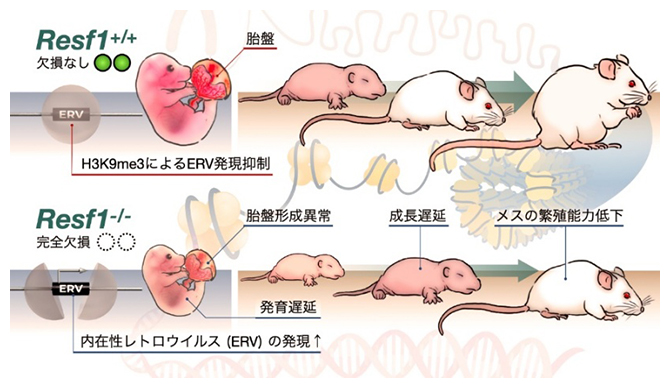
Resf1欠損で生じるマウスの異常
背景
生物の設計図ともいえる細胞のゲノムには、活性化すると転写やゲノム中の移動可能なトランスポゾン[1]をはじめとする多数の反復配列が存在します。反復配列の転写やゲノム中の移動は遺伝子発現の異常やゲノムの不安定化を引き起こします。そのため細胞は、反復配列をヘテロクロマチン[2]と呼ばれるクロマチン構造に収納することで転写やゲノム中の移動を抑制して、その活動を厳密に制御しています。ヘテロクロマチンの形成には、ヒストン(タンパク質)H3の9番目のリシン残基のメチル化(H3K9メチル化[3])が重要な役割を果たします。H3K9メチル化は、SETDB1やSUV39H1/2などのヒストンメチル化酵素によって担われ、トランスポゾンの転写抑制やゲノムの安定性維持に寄与しています。また、H3K9メチル化の分布は細胞種によって大きく異なり、特に胎盤由来細胞では数メガベース(数百万塩基対)に及ぶ大規模なヘテロクロマチン領域が形成されることが知られています。しかし、このような細胞種特有のヘテロクロマチンの形成や維持の機構はほとんど明らかになっていませんでした。
眞貝 主任研究員、福田 客員研究員はこれまでに、マウス胚性幹細胞(ES細胞)においてSETDB1依存的にトランスポゾンの発現を抑制する因子を網羅的に探索するスクリーニングを行い、その中からRESF1を新たな制御因子として発見しました注1)。しかし、RESF1が生体内でどのような役割を担い、個体発生やヘテロクロマチン形成にどのように関与しているのかは分かっていませんでした。
そこで本研究では、Resf1遺伝子を欠損させたマウスを作製し、生体内におけるRESF1の生理機能とヘテロクロマチン形成への役割を解析することに挑みました。
- 注1)2018年6月13日プレスリリース「ゲノム中のウイルスを抑制する仕組みの解明」
研究手法と成果
共同研究グループは、ES細胞においてSETDB1依存的なトランスポゾン抑制因子として同定していたRESF1の生理機能を明らかにするため、ゲノム編集技術を用いてResf1欠損マウスを作製しました。Resf1欠損マウスの出生率を解析したところ、ホモ接合体[4]はメンデルの法則(遺伝の法則)から予想される割合(25%)よりも有意に少なく、一部は胎生期に死亡することが分かりました。また、生存した個体においても体重減少や発育遅延が認められました。そこで胚発生を詳細に解析した結果、胎盤の重量の低下や構造異常など、顕著な構造異常が認められました。これらの結果から、RESF1は胎盤形成に重要な役割を果たしていることが明らかとなりました。次に胎盤における分子異常を解析したところ、トランスポゾンの発現上昇およびH3K9メチル化の変化が認められました(図1)。
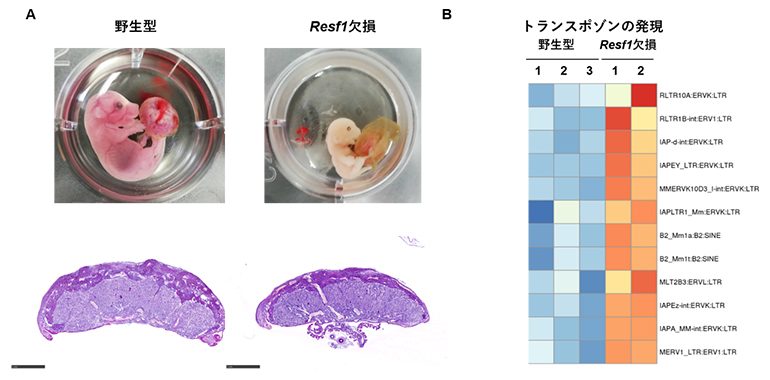
図1 Resf1欠損マウスにおける胎盤異常
- A.胎児致死および胎盤の形態異常。Resf1欠損マウスでは約半数が胎児期に致死となり(上段)、その個体では胎盤の著明な縮小が認められる(下段)。これらの結果は、Resf1が胎盤形成および胎児発生の維持に必須であることを示唆している。スケールバーは1mm。
- B.胎盤におけるトランスポゾン発現異常。胎盤組織におけるトランスポゾン発現を分子レベルで解析した結果、Resf1欠損胎盤では複数のトランスポゾンにおいて発現上昇(赤色)が認められた。この結果は、Resf1が胎盤においてトランスポゾン抑制機構の維持に関与している可能性を示している。
しかし胎盤は栄養膜幹細胞(TSC)[5]由来の複数の分化細胞種に加え、母体由来の免疫細胞や血管内皮細胞など、多様な細胞種から構成される組織です。そのため、組織レベルで観察されるエピゲノム(DNAの塩基配列そのものを変えることなくゲノムに加えられた修飾)変化が、どの細胞系譜に由来するのかを直接的に特定することは困難です。
そこで、Resf1欠損胚からTSCを樹立し、その分化系を用いて細胞種依存的な影響を1細胞RNAシークエンス[6]により解析しました。その結果、Resf1欠損TSCは自己複製能を維持しつつ、通常と同様に胎盤を構成する主要な細胞系譜へ分化しました。また、Resf1欠損は分化段階および細胞種ごとに異なる転写変動を引き起こすことが明らかとなりました。特に分化細胞において影響が顕著であり、RESF1が細胞種依存的に転写制御ネットワークを調節していることが示唆されました(図2)。
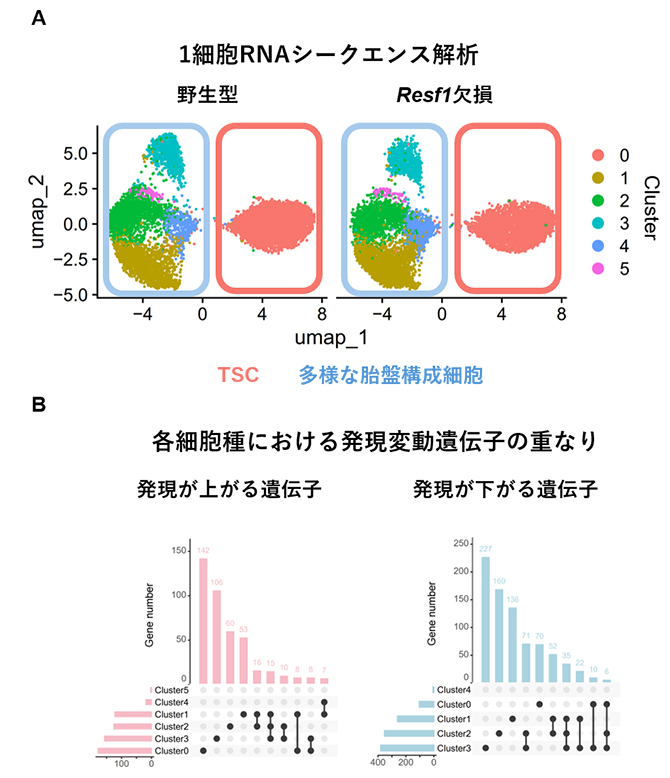
図2 TSCおよび分化TSCにおける遺伝子発現解析
- A.TSCおよび分化細胞の細胞構成。野生マウスおよびResf1欠損マウス由来TSCから分化させた細胞について1細胞RNAシークエンス解析を行い、各細胞の遺伝子発現プロファイルに基づいてUMAP解析(高次元データを2次元平面に投影する手法)を実施した。その結果、発現状態の類似性に応じて細胞集団が連続的に配置され、Resf1欠損細胞においてもTSC(オレンジ枠)から多様な分化細胞群(青枠)への分化軌跡が保持されていることが示された。
- B.細胞種特異的な遺伝子発現変動。各細胞種間でResf1欠損に伴う発現変動遺伝子を比較するため、サンプル間の発現変動遺伝子の重なりがどれくらいあるのかが一目で分かるUpSet Plot解析を行った。その結果、細胞種ごとに固有の発現変動遺伝子セットが多数存在することが明らかとなった。このことは、Resf1が一様な転写制御因子ではなく、細胞種依存的に異なる遺伝子群の発現制御に関与することを示唆している。
さらに、TSCおよび分化細胞におけるH3K9メチル化のChIP-seq[7]解析を行ったところ、トランスポゾン単体の制御変化に加えて、胎盤特異的な広域(メガベーススケール)のヘテロクロマチンドメイン(領域)においてH3K9メチル化の顕著な低下が認められました。この変化は特に分化細胞で顕著であり、RESF1が分化後のエピゲノム安定性維持に重要であることを示唆しています。この広域H3K9メチル化ドメインが従来知られているSETDB1依存的なヘテロクロマチン形成機構によって制御されているのかを検証するため、Setdb1欠損TSCにおけるH3K9メチル化を解析しました。その結果、Resf1欠損で観察された広域H3K9メチル化ドメインの低下はSetdb1欠損ではほとんど観察されず、両者でH3K9メチル化が低下する領域もほとんど重複しませんでした(図3)。
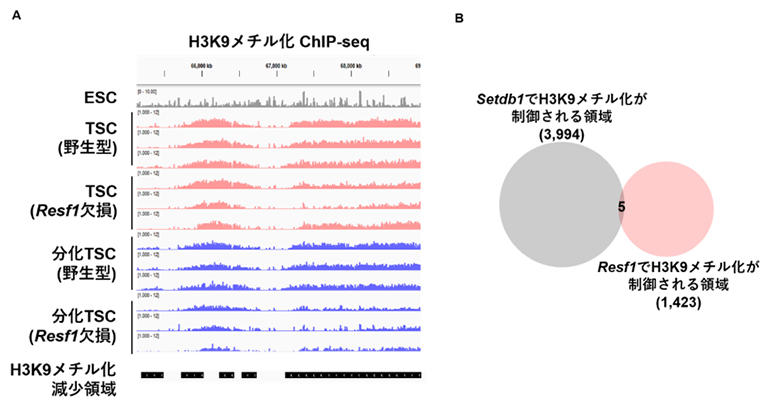
図3 Resf1で制御されるH3K9メチル化領域
- A.Resf1欠損におけるH3K9メチル化減少領域の例。ES細胞(ESC)ではH3K9メチル化は比較的小さなピークを形成するが、胎盤由来細胞では、メガベースレベルの大きなドメインを形成する。Resf1を欠損すると、このようなドメインのH3K9メチル化が減少する。
- B.TSCにおけるSETDB1とRESF1で制御されるH3K9メチル化領域の比較。両者はほとんど重ならないことから、TSCにおいてRESF1はSETDB1非依存的にH3K9メチル化を制御することが示唆される。
これらの結果から、RESF1は従来考えられていたSETDB1依存的なヘテロクロマチン形成経路とは異なる機構を介して、胎盤細胞特異的な広域ヘテロクロマチンドメインの維持に寄与していることが明らかとなりました。そこで、RESF1が協調して機能する因子を探索したところ、RESF1は別のヒストンメチル基転移酵素SUV39H1/2と相互作用する可能性が見いだされました。SUV39H1/2は遺伝子密度の低い領域に広域H3K9メチル化ドメインを形成することが知られていることから、RESF1はSUV39H1/2と協調して胎盤細胞特異的なヘテロクロマチン構造を維持しているのではと推察されます。
以上のように、トランスポゾン抑制因子として同定されていたRESF1は、生体内においてもトランスポゾン抑制に機能するとともに、胎盤発生を含む胚発生過程に重要な役割を果たすことが明らかとなりました。さらに、RESF1は従来想定されていたSETDB1依存経路とは異なる機構を介して、細胞種特異的な広域ヘテロクロマチン状態を維持することが明らかとなりました。
今後の期待
本研究により、RESF1はトランスポゾン抑制因子として機能するだけでなく、細胞種特異的なヘテロクロマチン形成を介して胎盤発生を制御する新たなエピゲノム制御因子であることが明らかとなりました。一方で、RESF1がSUV39H1/2とどのように協調して広域ヘテロクロマチンドメインを形成・維持するのか、その分子機構の全容は未解明です。今後は、RESF1複合体の構成因子やクロマチンへのリクルート機構を明らかにすることで、細胞種ごとに異なるヘテロクロマチン制御機構の理解がさらに進むことが期待されます。また、本研究で見いだされた胎盤特異的なエピゲノム制御機構は、胎盤形成異常や胎児発育不全などの発生異常の理解に加え、トランスポゾン制御異常が関与するさまざまな疾患の分子基盤の解明にもつながることが期待されます。
補足説明
- 1.トランスポゾン
ゲノム中を移動(転移)できるDNA配列の総称で、宿主遺伝子の転写調節やゲノム不安定性の原因にもなる。 - 2.ヘテロクロマチン
高度に凝縮したクロマチン領域で、転写が抑制されやすく、反復配列やトランスポゾンが多く含まれるゲノム領域。 - 3.H3K9メチル化
ヒストンH3の9番目リジン残基のメチル化修飾で、ヘテロクロマチン形成とトランスポゾン抑制に関わる代表的なエピジェネティック修飾。 - 4.ホモ接合体
生物のある遺伝子座において、両親から受け継いだ遺伝子が同一である状態を指す。 - 5.栄養膜幹細胞(TSC)
胚発生初期の栄養膜系譜を維持し、胎盤形成に寄与する幹細胞で、増殖能と分化能を併せ持つ細胞集団。TSCはtrophoblast stem cellsの略。 - 6.1細胞RNAシークエンス
個々の細胞ごとのトランスクリプトーム(転写産物全体)を解析する手法で、細胞集団の不均一性や分化状態を高解像度で解析できる技術。 - 7.ChIP-seq
DNAと結合しているタンパク質(転写因子やヒストン修飾)を免疫沈降で回収し、その結合領域をゲノムワイドに同定する解析手法。
共同研究グループ
理化学研究所
開拓研究所
眞貝細胞記憶研究室
客員研究員 福田 渓(フクダ・ケイ)
(山梨大学 大学院生命環境学域 器官形成ダイナミクス研究室 助教)
主任研究員 眞貝 洋一(シンカイ・ヨウイチ)
テクニカルスタッフⅠ 志村 知古(シムラ・チカコ)
バイオリソース研究センター
統合発生工学研究開発室
研究員 小倉 淳郎(オグラ・アツオ)
室長 井上 貴美子(イノウエ・キミコ)
専任研究員 的場 章悟(マトバ・ショウゴ)
テクニカルスタッフⅡ 廣瀬 美智子 (ヒロセ・ミチコ)
山梨大学 生命環境学域
発生ゲノム科学研究室
助教 北澤 萌恵(キタザワ・モエ)
細胞ポテンシャル研究室
准教授 石内 崇士(イシウチ・タカシ)
研究支援
本研究は、理研新領域開拓課題「Genome building from TADs/ゲノム構築原理の理解に向けて(研究代表者:眞貝洋一、小倉淳郎)」により実施し、日本学術振興会(JSPS)科学研究費助成事業基盤研究(A)「ヘテロクロマチン形成・維持の分子機構の解明(研究代表者:眞貝洋一、JP22H00413)」、同新学術領域研究(研究領域提案型)「細胞分化にともなうクロマチンポテンシャルの変化とその分子基盤(研究代表者:眞貝洋一、JP18H05530)」、同若手研究「高次クロマチンのマクロ構造の制御機構の解明(研究代表者:福田渓、JP22K15044)」、同挑戦的研究(開拓)「naive型胎盤幹細胞(TS細胞)の開発とその応用研究(研究代表者:眞貝洋一、JP24K21266)」、同学術変革領域研究(A)「生殖工学技術による生殖ライフスパンの操作(研究代表者:眞貝洋一、JP23H04956)」、科学技術振興機構(JST)創発的研究支援事業「胎盤による胚発生の保護メカニズムとその破綻(研究代表者:的場章悟、JPMJFR221G)」による助成を受けて行われました。
原論文情報
- Kei Fukuda, Kimiko Inoue, Chikako Shimura, Moe Kitazawa, Michiko Hirose, Takashi Ishiuchi, Shogo Matoba, Atsuo Ogura, Yoichi Shinkai, "Resf1 is required for proper placental development and configuration of trophoblast cell-specific heterochromatin", Genome Research, 10.1101/gr.281507.125
発表者
理化学研究所
開拓研究所 眞貝細胞記憶研究室
客員研究員 福田 渓(フクダ・ケイ)
テクニカルスタッフⅠ 志村 知古(シムラ・チカコ)
主任研究員 眞貝 洋一(シンカイ・ヨウイチ)
 福田 渓
福田 渓
 眞貝 洋一
眞貝 洋一
報道担当
理化学研究所 広報部 報道担当
お問い合わせフォーム