2026年7月16日
理化学研究所
名古屋大学
ウィタヤシリメティー科学技術大学院大学
岡山大学
科学技術振興機構(JST)
ヒト細胞骨格の“祖先”を古細菌で発見
-微小管の進化の謎に新知見-
理化学研究所(理研)生命医科学研究センター 構成的細胞生物学研究チームのツアン・トゥイ・リン 特別研究員、宮﨑 牧人 チームディレクター(科学技術振興機構(JST)創発研究者)、名古屋大学 大学院理学研究科 生命理学専攻の成田 哲博 准教授、ウィタヤシリメティー科学技術大学院大学(VISTEC)生体分子理工学研究科のロビンソン・チャールズ・ロバート 教授(岡山大学 学術研究院先鋭研究領域(異分野基礎科学研究所)客員教授)らの国際共同研究グループは、約27億年前に誕生したと考えられるアスガルド古細菌[1]が持つタンパク質を分析し、真核生物の細胞骨格の祖先と考えられる「原始的な微小管[2]」を発見しました。
本成果は、微小管の進化の過程を理解する上で重要な知見であり、真核生物誕生の謎の解明に貢献すると期待されます。
微小管はチューブリン(tubulin)[3]というタンパク質がたくさん集まる重合という反応でできた、管状の線維です。私たちヒトを含め真核生物に共通する重要な細胞内構造で、細胞の形の制御や細胞分裂、細胞内の物質輸送などの重要な機能を担っています。これまで、複雑な微小管がどのように進化したのかはよく分かっていませんでした。今回、国際共同研究グループは、真核生物に最も近縁とされるアスガルド古細菌の一種である、ヘイムダル古細菌が持つチューブリン様タンパク質の構造と機能を調べました。その結果、真核生物の微小管に特徴的な四つの性質を持ちながらも、真核生物よりも細くて単純な"ミニマル(最小限の)微小管"をつくることを発見しました。
本研究は、科学雑誌『Science Advances』オンライン版(7月15日付:日本時間7月16日)に掲載されました。
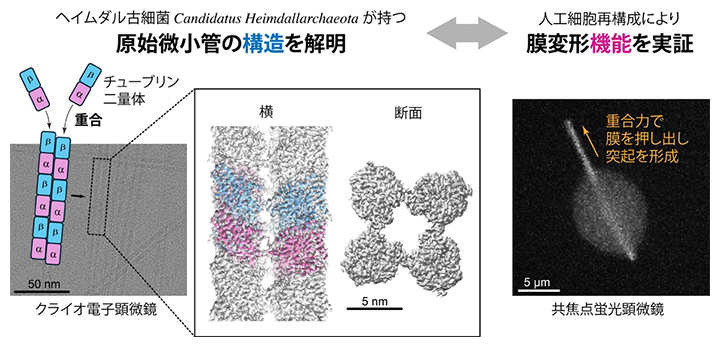
今回の研究で明らかにした原始的な微小管の構造と機能
背景
私たちの体をつくる真核細胞の内部には、「細胞骨格」と呼ばれる線維状の構造があります。その中でも微小管は、細胞の形を支えたり、細胞分裂のときに染色体を分けたり、細胞内で物質を運ぶ"レール"として働いたりする重要な構造です。微小管は、αチューブリンとβチューブリンという二つのタンパク質が組み合わさってできる管状の線維です。真核生物の微小管は通常、13本の細いプロトフィラメント[4]と呼ばれる線維がリング状に集まり、太くて硬い管をつくります。この構造により、微小管は細胞の中で長い距離を伸び、押す力に耐えることができます。一方で、微小管はただ安定な棒ではなく、伸びたり急に縮んだりする「動的不安定性(ダイナミックインスタビリティー)[5]」と呼ばれる性質を持っています。この動きは、細胞分裂や細胞内輸送に欠かせません。しかし、このような複雑な微小管が、進化の過程でどのように生まれたのかはよく分かっていませんでした。
近年、真核生物に最も近い原核生物とされるアスガルド古細菌(図1)の一部にも、チューブリンに似たタンパク質が見つかってきました。これらのタンパク質は、真核細胞の微小管の起源を考える上で重要な手掛かりになります。しかし、多くのアスガルド古細菌は培養が非常に難しく、細胞内での働きを直接調べることが困難です。
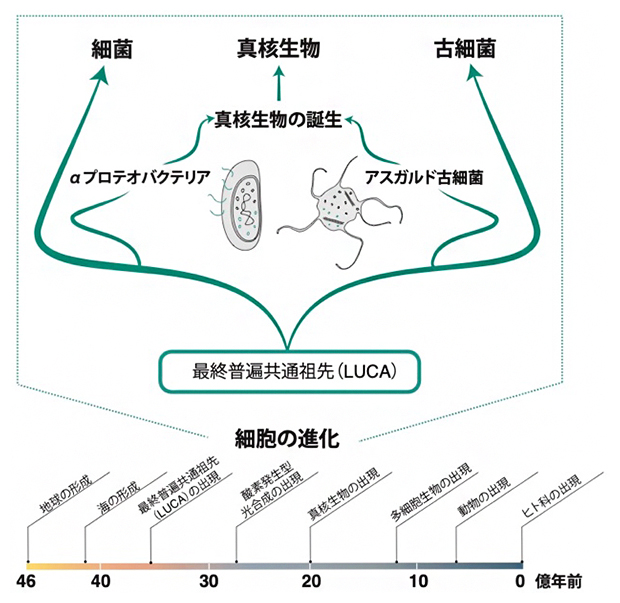
図1 細胞進化の概略図
最終普遍共通祖先(LUCA)から細菌と古細菌が分岐し、アスガルド古細菌とαプロテオバクテリアの共生を経て、約20億年前に真核生物が誕生したと考えられている。
研究手法と成果
今回、国際共同研究グループは、未培養のアスガルド古細菌の一種、ヘイムダル古細菌(Candidatus Heimdallarchaeota)が持つ二つのチューブリン様タンパク質に着目しました。これらのタンパク質の遺伝子を大腸菌に組み込み、大腸菌にタンパク質をつくらせた後、それらを精製して試験管内や人工細胞に封入して機能を調べるという方法を採りました。そして、これらが真核生物のようにαチューブリンとβチューブリンが結合したα/βチューブリン二量体をつくるのか、微小管のような管をつくるのか、さらに伸び縮みや力の発生といった機能を持つのかを調べました。その結果、ヘイムダル古細菌由来のチューブリンは、真核生物の微小管で重要な四つの機能と共通する基本的な性質を持ちながら、より小さな4本構造の微小管をつくることが分かりました。
まず、米国サンフランシスコ近郊の河口域の泥(表層堆積物)から得られたメタゲノム情報(特定環境中の核酸配列を網羅的に解析して得られた情報)から、ヘイムダル古細菌が持つ二つのチューブリン様遺伝子を同定しました(図2A)。この二つの遺伝子はゲノム上で隣り合っており、同じ向きに並んでいました。これは、二つのタンパク質が細胞内で同時につくられ、協調して働く可能性を示しています。進化の系統を推定する系統解析の結果、これらのタンパク質は、真核生物のαチューブリン、βチューブリンに対応するような二つのグループに分かれることが分かりました。
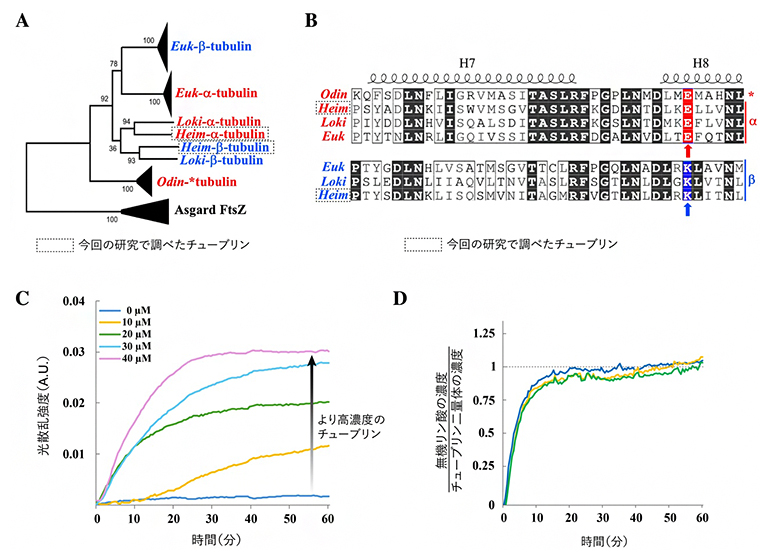
図2 チューブリンの生物種間比較とHeimチューブリンの生化学分析
- A真核生物(Euk)および3種類のアスガルド古細菌(Loki、Odin、Heim)が持つチューブリンの系統樹。アミノ酸配列を比較し、進化の過程として最も起こりやすい系統関係を統計的に推定する最尤法(さいゆうほう)を用いて作成した。外群として原核生物に共通するチューブリンに似たタンパク質(FtsZ)の配列を用いた。赤色と青色は、それぞれGTPの触媒活性を持つグルタミン酸残基、および触媒活性を持たないリシン残基を有するチューブリンを示す。
- BGTPが結合する部位(H7およびH8ヘリックス)のアミノ酸配列の比較。赤色および青色の矢印は、それぞれグルタミン酸残基およびリシン残基を示す。
- C濃度依存的なHeim-α/βチューブリン二量体の重合反応。GTP存在下、37℃で反応を行った。縦軸の値が大きいほど、よりたくさんの二量体が集まって複合体が形成されたことを示す。A.U.:任意単位(arbitrary unit)。µM(マイクロモーラー、1マイクロモーラーは100万分の1モル/L)。
- D重合中のGTPの分解反応。GTPが分解されたときに産生される無機リン酸を測定した。無機リン酸の濃度のHeim-α/βチューブリン二量体濃度に対する比(縦軸の値)が、おおよそ1に収束したことから、重合には二量体当たり1個のGTPの分解が必要と分かった。3回の独立実験データを示す。
そこで国際共同研究グループは、それぞれをHeim-αチューブリン、Heim-βチューブリンと名付けました。さらに配列を詳しく調べると、一方にはGTP(グアノシン三リン酸)[6]加水分解に関わるグルタミン酸が、もう一方には加水分解を起こしにくくするリシンが保存されていました(図2B)。これは、真核生物のαチューブリンとβチューブリンに似た役割分担が、Heim-αチューブリンとHeim-βチューブリンにもあることを示唆しています。
次に、Heim-αチューブリンとHeim-βチューブリンを大腸菌で発現させ、精製しました。二つを同時に発現させると、Heim-αチューブリンとHeim-βチューブリンが自然に結合して、安定なHeim-α/βチューブリン二量体として精製できました。一方、Heim-βチューブリン単独では不安定で変性しやすいことが分かり、Heim-αチューブリンとの組み合わせが安定性に重要であることが示されました。
精製したHeim-α/βチューブリン二量体にGTPを加えて37℃で温めると、タンパク質濃度に応じて重合が進みました(図2C)。一方、GTPの代わりにGDP(グアノシン二リン酸)[6]を加えた場合には、ほとんど重合は起こりませんでした。このことから、Heim-α/βチューブリン二量体の重合はGTPに依存することが分かりました。さらにGTPが分解されたときに産生される無機リン酸の測定から、Heim-αあるいはHeim-βチューブリンのいずれかによって、1個のGTPが分解されたことが判明しました(図2D)。この性質は、真核生物の微小管とよく似ています。
次に、クライオ電子顕微鏡[7]を用いて、Heim-α/βチューブリン二量体がつくる構造(Heim微小管)を観察しました(図3A)。80万本の線維状構造の画像に平均化処理を施して詳細な構造を解析したところ、これらの線維は4本のプロトフィラメントから成る管状構造であること(図3B)、管状構造には2種類あることが分かりました(図3C)。いずれの構造も、真核生物の一般的な13本のプロトフィラメント微小管よりも小さな構造です。
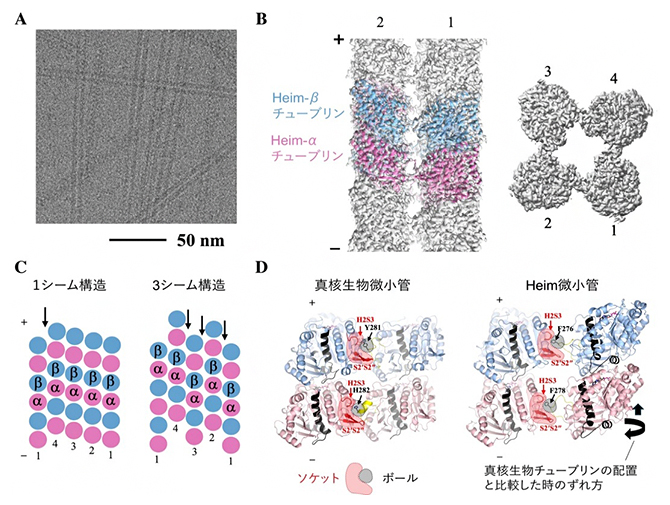
図3 Heim微小管の構造
- AHeim微小管のクライオ電子顕微鏡画像。スケールバーは50ナノメートル(nm、1nmは10億分の1メートル)。
- B電子顕微鏡画像の解析で得られたHeim微小管構造。左:上から見た図。右:横から見た図。2種類の構造のうち、主要な方の構造を示す(詳細は(C)参照)。+および-は微小管の極性を示す。
- C発見した2種類の微小管構造の模式図。プロトフィラメント間にシームと呼ばれるずれが見られた。左:1シーム構造(真核生物微小管と同じシーム数)。右:3シーム構造。1シーム構造の割合の方が多かったことから、細胞内では主に1シーム構造を取ると考えられる。
- D真核生物微小管(左)とHeim微小管(右)のプロトフィラメント間相互作用の比較。ボールとソケットの相互作用の仕方は両者で保存されていることが分かる。
国際共同研究グループは、高分解能の構造解析を行い、主要な構造について2.8オングストローム(1Å、1Åは100億分の1メートル)の空間分解能を達成しました。その結果、隣り合うプロトフィラメント同士が、真核生物の微小管にも見られるボールとソケットのような相互作用で結合していることが明らかになりました(図3D)。つまり、管の太さは大きく異なるものの、微小管を安定化する基本的な結合様式は保存されていました。この結果は、チューブリンがつくる管状構造の基本原理が、真核生物が成立する以前から存在していた可能性を示しています。
さらに、蛍光顕微鏡を用いて、1本1本のHeim微小管がどのように伸び縮みするかを観察しました(図4A)。安定化した短いHeim微小管を種としてガラス表面に固定し、別の色で蛍光標識したHeim-α/βチューブリン二量体を加えると、微小管は両端から伸長しました。ただし、二つの端の伸長速度は同じではなく、有意な差を示しました。これはHeim微小管が真核生物の微小管と同じように「両端で重合の性質が異なる極性」を持つことを示しています(図4B)。
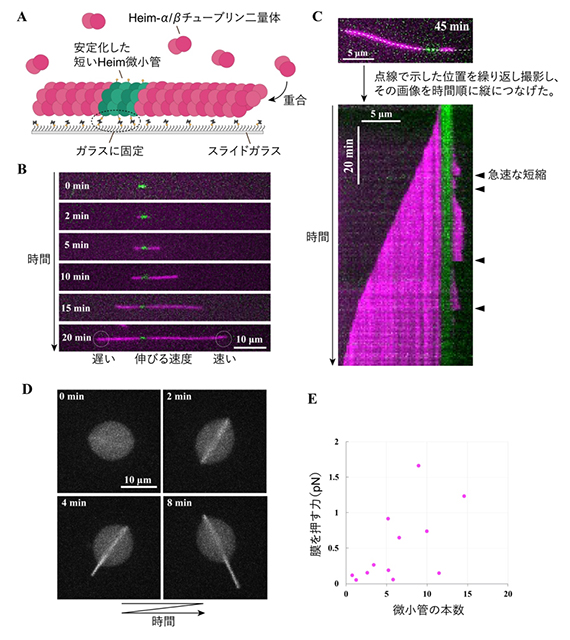
図4 蛍光顕微鏡を用いたHeim微小管の重合過程と膜変形過程の観察
- A実験方法。スライドガラスを横から見た模式図。安定化した短いHeim微小管を種としてスライドガラス表面に固定し、別の色で蛍光標識したHeim-α/βチューブリン二量体を加えると、種微小管から伸びる様子が観察できる。
- BHeim-α/βチューブリン二量体(マゼンタ)が、種のHeim微小管(緑色)に結合して伸びる様子。
- CHeim微小管が示す動的不安定性。片方の端では、伸びた後に急激に縮む現象が繰り返し生じた。
- DHeim-α/βチューブリン二量体を封入したリポソームの顕微鏡画像。リポソーム内でHeim微小管が形成され、それらが束になって膜を押すことで突出構造が形成された。
- EHeim微小管の束が膜を押す力。個々のリポソームの測定結果をプロットした。平均で6本の微小管が束となり、束全体で0.5pNの力を生み出すことが分かった。
チューブリン濃度を中間的な濃度に下げると、一方の端では、伸長と急速な短縮が確率的に切り替わる様子が観察されました。これは、真核生物の微小管に特徴的な「動的不安定性」とよく似ています。特に低濃度の条件下では、片方の端がのこぎり歯状の長さ変化を示し、伸びた後に急激に縮む現象が明瞭に観察されました。一方、反対側の端では同様の現象は確認されませんでした(図4C)。このことから、Heim微小管にも、真核生物の微小管と同じようにプラス端とマイナス端に相当する機能的な違いがあることが分かりました。
最後に、Heim微小管が膜を変形させる力を生み出せるかを調べるため、国際共同研究グループは、精製したHeim-α/βチューブリン二量体をリポソーム[8]の中に封入しました。リポソームに封入してから温度を上げて重合反応を開始すると、内部でHeim微小管が形成され、複数のHeim微小管が束をつくる様子が観察されました。直径10~20マイクロメートル(μm、1μmは100万分の1メートル)のリポソームでは、70%以上のリポソームで明瞭な形の変化が起こりました。その多くでは、リング状の束によってリポソームが楕円形(だえんけい)に変形しました。さらに小さなリポソームでは、35%のリポソームで膜が外側に突き出す安定な突出構造が形成されました(図4D)。これは、Heim微小管の伸長が膜を押す力を生み出せることを示しています。
力学的性質を調べたところ、Heim微小管の持続長[9]は、おおよそ500μmでした。これは真核生物が持つ主要な細胞骨格であるアクチン線維の持続長よりも約30倍長く、Heim微小管はかなり硬い線維であることを示しています。一方で、真核生物の微小管の持続長は1~10mm程度であり、Heim微小管はそれより柔らかい構造でした。
さらに、膜突出時に観察されたHeim微小管の束が、力がかかるとある段階で急激に曲がる座屈(ざくつ)という現象から、Heim微小管が発生する押す力を推定しました。束全体では平均0.5ピコニュートン(pN、1pNは1兆分の1ニュートン)の力が見積もられました(図4E)。1本のHeim微小管当たりの力は平均で0.08pNでした。これは真核生物のアクチン線維が生み出す力と同程度ですが、真核生物の微小管で報告されている3~4pNよりは小さい値です。
以上の結果から、Heim-α/βチューブリンニ量体はGTP依存的に重合し、4本のプロトフィラメントから成る小型の微小管をつくり、極性と動的不安定性を示し、さらに膜を変形させる力を生み出せることが明らかになりました。これは、真核生物の微小管が持つ主要な性質が、より原始的な生物のチューブリンシステムにも備わっていることを示しています。
今後の期待
今回の研究は、真核細胞に特徴的な微小管が、どのような進化的過程を経て現在の13本のプロトフィラメント構造になったのかを考える新しい手掛かりを与えます。Heim微小管は4本のプロトフィラメントから成る小型の構造ですが、「GTP依存的重合」、「両端で重合の性質が異なる極性」、「動的不安定性」、「重合による力発生」という、真核生物の微小管に特徴的な四つの性質を備えていました。
このことは、微小管の機能の核となる仕組みが、必ずしも13本のプロトフィラメント構造を必要としないことを示しています。一方で、Heim微小管は真核生物の微小管よりも柔らかく、発生する力も小さいことが分かりました。この違いは、細胞が大型化し、鞭毛(べんもう)運動や複雑な細胞分裂装置を進化させる過程で、より太くて硬い13本のプロトフィラメント微小管が選ばれた可能性を示しています。小さな細胞では、少ないタンパク質量で細胞内を横断できる4本のプロトフィラメント構造が有利だった可能性があります。一方、大きな細胞では、より強い力に耐えられる太い微小管が必要になったと考えられます。この成果は、細胞骨格の進化を理解する基礎研究として重要であるだけでなく、われわれヒトを含む真核生物が誕生した謎の解明への重要な知見となります。
今回、少数のタンパク質だけで膜を変形させる仕組みを人工的に再現した点は、人工細胞や合成生物学への応用にもつながります。将来的には、微小管の太さや硬さを設計することで、人工細胞の形を制御したり、膜を押して変形させる分子機械をつくったりする研究へ発展する可能性があります。さらに、今回の成果は、細胞の形や動きを分子から設計するための基盤技術になると期待されます。
補足説明
- 1.アスガルド古細菌
真核生物に最も近縁と考えられている古細菌のグループ。真核細胞の起源を考える上で重要な生物群であることから、近年注目を集めている。 - 2.微小管
真核細胞に備わる細胞骨格の一種。チューブリン([3]参照)というタンパク質からできた管状の線維で、細胞の形の維持、細胞分裂、細胞内輸送などに関わる。 - 3.チューブリン(tubulin)
微小管をつくるタンパク質。αチューブリンとβチューブリンが二量体をつくり、それが多数集まって微小管を形成する。 - 4.プロトフィラメント
チューブリンが縦方向につながった細い線維。真核生物の一般的な微小管では、13本のプロトフィラメントが横に並んで管をつくる。 - 5.動的不安定性(ダイナミックインスタビリティー)
微小管が伸長と急速な短縮を確率的に切り替える性質。細胞分裂時に、微小管が染色体を効率的に探索するために重要であると考えられている。 - 6.GTP(グアノシン三リン酸)、GDP(グアノシン二リン酸)
GTPは細胞内でエネルギーや分子スイッチとして働く物質。チューブリンの重合や微小管の動的不安定性に関わる。GDPは、GTPがエネルギーを放出した後の状態の分子である。真核生物α/βチューブリン二量体は、αチューブリンとβチューブリンのそれぞれがGTPを結合した状態で微小管に取り込まれ、その後βチューブリンに結合しているGTPのみがGDPへと変換(加水分解)される。この変換により微小管は構造が不安定化し、崩壊が促進されるため、GTPからGDPへの変換が微小管の成長と崩壊を制御する重要な仕組みと考えられている。 - 7.クライオ電子顕微鏡
タンパク質などの生体分子を水溶液中の生理的な環境に近い状態で、電子顕微鏡で観察するために開発された手法。試料を含む水溶液を急速凍結して、液体窒素温度(-196℃)付近まで冷却することで電子線の照射による損傷を低減し、生体分子やその複合体の構造解析を行うことができる。 - 8.リポソーム
脂質膜でできた人工的な小胞。細胞膜を単純化したモデルとして、細胞内構造の再構成実験や人工細胞の合成に用いられる。 - 9.持続長
線維の硬さを表す指標。値が大きいほど曲がりにくく、硬い線維であることを示す。
国際共同研究グループ
理化学研究所 生命医科学研究センター 構成的細胞生物学研究チーム
チームディレクター 宮﨑 牧人(ミヤザキ・マキト)
(生命機能科学研究センター 上級研究員、JST創発研究者)
特別研究員 ツアン・トゥイ・リン(Linh Thuy Tran)
テクニカルスタッフⅠ 山崎 陽祐(ヤマザキ・ヨウスケ)
名古屋大学 大学院理学研究科 生命理学専攻
准教授 成田 哲博(ナリタ・アキヒロ)
研究員 松本 友治(マツモト・トモハル)
岡山大学 学術研究院先鋭研究領域(異分野基礎科学研究所)
助教(特任)アリ・サムソン(Samson Ali)
ウィタヤシリメティー科学技術大学院大学(VISTEC)(タイ)生体分子理工学研究科
教授 ロビンソン・チャールズ・ロバート(Robert Charles Robinson)
(岡山大学 学術研究院先鋭研究領域(異分野基礎科学研究所)客員教授)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費助成事業若手研究「A study on microtubule evolution from Asgard OdinTubulin(研究代表者:Linh Thuy Tran、JP25K18436)」、同学術変革領域研究(A)「生体内力作用の定量的計測技術開発(研究代表者:吉村成弘、研究分担者:宮﨑牧人、JP22H05171)」、同基盤研究(B)「疎らに結合したアクチン結合タンパク質がアクチン線維全体の機能を調節するメカニズム(研究代表者:上田太郎、研究分担者:成田哲博、JP23K27145)」「微小管の進化再現とそれによる微小管動態の理解(研究代表者:成田哲博、JP26K01973)」、同基盤研究(A)「Eukaryotic-like proteins from Asgard archaea(研究代表者:Robert Charles Robinson、JP20H00476)」、理研奨励研究プロジェクト「Structural and functional analysis of Heimdallarchaeaota tubulin reveals the origin of eukaryotic microtubules(研究代表者:Linh Thuy Tran)」、同生命機能科学研究センター構造細胞生物学プロジェクト(研究代表者:宮﨑牧人)、笹川科学研究助成「ヘイムダル古細菌チューブリンの構造機能解析から迫る真核生物微小管の誕生起源(研究代表者:Linh Thuy Tran、2025-4021)」、文部科学省科学技術人材育成費補助事業「ダイバーシティ研究環境実現イニシアティブ(研究代表者:Linh Thuy Tran、500230057)」、科学技術振興機構(JST)創発的研究支援事業「再構成で紐解く細胞骨格機能の自己組織化メカニズム(研究代表者:宮﨑牧人、JPMJFR2417)」、同戦略的創造研究推進事業 CREST「合成細菌JCVI syn3.0Bとゲノム操作を用いた細胞進化モデルの構築(研究代表者:宮田真人、研究分担者:Robert Charles Robinson、JPMJCR19S5)」、Federation of European Microbiological Societies(FEMS)「Moore-Simons Project on the Origin of the Eukaryotic Cell(研究代表者:Robert Charles Robinson、GBMF9743)」、Nagoya University Research Fundの助成を受けて行われました。クライオ電子顕微鏡を用いた構造解析は、岡山大学異分野基礎科学研究所の国際構造生物学研究センターの支援の下で実施されました。
原論文情報
- Linh T. Tran, Samson Ali, Tomoharu Matsumoto, Yosuke Yamazaki, Akihiro Narita, Makito Miyazaki, Robert C. Robinson, "Structure and dynamics of a four-protofilament microtubule from Heimdallarchaeales α/β -tubulin", ChemBioChem, 10.1002/cbic.201800620
発表者
理化学研究所
生命医科学研究センター 構成的細胞生物学研究チーム
特別研究員 ツアン・トゥイ・リン(Linh Thuy Tran)
チームディレクター 宮﨑 牧人(ミヤザキ・マキト)
(JST創発研究者)
名古屋大学 大学院理学研究科 生命理学専攻
准教授 成田 哲博(ナリタ・アキヒロ)
ウィタヤシリメティー科学技術大学院大学(VISTEC)(タイ)生体分子理工学研究科
教授 ロビンソン・チャールズ・ロバート(Robert Charles Robinson)
(岡山大学 学術研究院先鋭研究領域(異分野基礎科学研究所)客員教授)
発表者のコメント
It would be an honor to continue my research at RIKEN and to collaborate with other labs in Japan, sharing ideas and publishing our work together. Through step-by-step investigations into microtubule evolution using in vitro assays and structural biology, we hope our research will contribute to the cytoskeleton field and benefit those interested in understanding eukaryotic-like proteins encoded in Asgard archaeal genomes.(Linh Thuy Tran)
生命の進化の痕跡を目の当たりにし、生命のロマンを垣間見ることができたことに、とても興奮しました。新しい結果が次々と得られたときは、研究者を続けていて良かったと感じる、幸せな瞬間でした。この研究は、異なる専門性を持つ共同研究者の力があって初めて実現しました。異分野の研究者と協力することで、自分一人では思いつかなかった視点や発見にたどり着くことができたことにも、大きな喜びを感じています。(宮﨑 牧人)
JST事業に関する問い合わせ
科学技術振興機構 創発的研究推進部
東出学信(ヒガシデ・タカノブ)
Tel: 03-5214-7276
Email: souhatsu-inquiry@jst.go.jp
報道担当
理化学研究所 広報部 報道担当
お問い合わせフォーム
名古屋大学 総務部広報課
Tel: 052-558-9735
Email: nu_research@t.mail.nagoya-u.ac.jp
ウィタヤシリメティー科学技術大学院大学
Tel: + 66-33-014-444
Email: Ratthanan.b@vistec.ac.th(Ratthanan Butrsikhao)
岡山大学 総務部 広報課
Tel: 086-251-7292
Email: www-adm@adm.okayama-u.ac.jp
科学技術振興機構 広報課
Tel: 03-5214-8404
Email: jstkoho@jst.go.jp