理化学研究所(理研)環境資源科学研究センター 細胞機能研究チームのダンカン・コールマン 特別研究員(研究当時、現 客員研究員)、杉本 慶子 チームディレクターらの国際共同研究グループは、傷害が植物の再生を誘導する新たな仕組みを発見しました。本研究成果は、植物の再生の理解を深めるとともに、穀物や野菜、果樹などにおける再生・育種技術の高度化に貢献すると期待されます。
植物は動物の体と同じように、さまざまな役割を持った細胞が集まってできています。こうした植物では、気候条件や害虫による被害を受けて傷つくと、その周辺の細胞が担っていた役割(分化状態)を変える「細胞リプログラミング[1]」によって、葉や茎などの器官を再生する能力を持っています。この高い再生能力は植物の特徴の一つですが、傷害という環境刺激がどのように細胞リプログラミングを引き起こすのか、その分子メカニズムは解明されていませんでした。
今回、国際共同研究グループは、主に猛暑などの高温環境にさらされた際に生じる熱ストレス応答[2]に関わる転写因子HSFA1[3]が、傷害に応答して細胞リプログラミングを誘導する中心的な役割を担うことを明らかにしました。さらに、HSFA1はWIND1[4]、PLT3[5]、ZAT6[6]などのリプログラミング関連因子を直接活性化することが分かりました。これにより、傷害シグナルがどのように転写ネットワーク[7]を立ち上げ、細胞のリプログラミングを引き起こすのかが明らかになりました。本研究は、熱ストレス応答因子として知られてきたHSFA1が植物再生の制御において中核的に機能することを示し、環境応答と発生制御を結び付ける新たな分子基盤を提示するものです。本研究は、科学雑誌『The Plant Cell』オンライン版(4月22日付)に掲載されました。
背景
植物は傷害を受けた際に失われた器官を再生する優れた能力を持っており、この能力は農業やバイオテクノロジー分野における重要な基盤となっています。例えば、作物の栽培において、台風や害虫による被害を受けた植物が自己修復する能力や、挿し木による増殖技術は、この再生能力に基づいています。植物は、傷口周辺の細胞が分化状態を変化させる「細胞リプログラミング」という現象を通じて茎葉(けいよう)や根などの器官を再生できます。
杉本チームディレクターらは、これまでに植物が新たな器官を再生する仕組みについて研究を進め、植物の細胞リプログラミングを制御する機構を明らかにしてきました注1~8)。これまでの研究により、WIND1やそのホモログ(相同体)[8]、PLT3やそのホモログなどの細胞リプログラミングを促進する因子が同定され、その制御機構の一部が明らかにされてきました。しかしながら、傷害という環境刺激がどのようにしてこれらの因子を活性化させ、細胞リプログラミングを引き起こすのか、「傷害シグナル」と「遺伝子発現制御」とをつなぐ上流の分子機構については、依然として多くが未解明のままでした。
熱ストレスが加わった直後に応答するマスター制御因子のHSFA1は、熱ストレスに応答して核内へ移行し、下流で働く他のHSFホモログ遺伝子や細胞を保護する働きを担うHSP遺伝子発現を誘導することが知られていましたが、傷害応答における役割は不明でした。
今回、国際共同研究グループは、シロイヌナズナを用いてHSFA1の植物再生の制御における機能を検証することにより、傷害応答における初期の転写制御機構の解明を目指しました。
- 注1)2011年3月11日プレスリリース「植物細胞の脱分化を促進するスイッチ因子を発見」
- 注2)2015年6月30日プレスリリース「植物の分化全能性抑制の分子メカニズムの一端を解明」
- 注3)2017年1月17日プレスリリース「植物が傷口で茎葉を再生させる仕組み」
- 注4)2019年11月4日プレスリリース「植物細胞のリプログラミングの仕組みを分子レベルで解明」
- 注5)2021年8月20日プレスリリース「植物の再生と防御のスイッチ」
- 注6)2022年8月4日プレスリリース「分化細胞からの植物体再生」
- 注7)2024年9月27日プレスリリース「光が植物の再生運命を決める」
- 注8)2026年3月24日プレスリリース「植物再生スイッチの制御原理を解明」
研究手法と成果
まず、RNA次世代シーケンス(RNA-seq)[9]データを再解析し、傷害を受けた直後に発現が高まる遺伝子群を同定しました。結果、その遺伝子群にはHSFA1の下流因子として知られるHSFA2やHSFA7A遺伝子をはじめ、熱ストレスによって誘導される遺伝子群が多数含まれていることが分かりました(図1)。
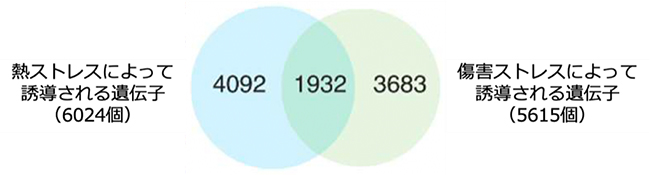
図1 熱ストレスと傷害ストレスに共通して誘導される遺伝子の同定
公開RNA-seqデータの再解析により、傷害ストレス後12時間以内に発現上昇する5,615個の遺伝子のうち、1,932個の遺伝子が熱ストレスによっても誘導されることが明らかになった。
次に、HSFA1が、熱ストレスへの応答時と同様に、傷害に応答して核内へ移行するのかを検証しました。その結果、傷害を与える前には細胞質に局在していたHSFA1タンパク質が、傷害を与えて15分後には核内へ移行することが明らかになりました(図2)。
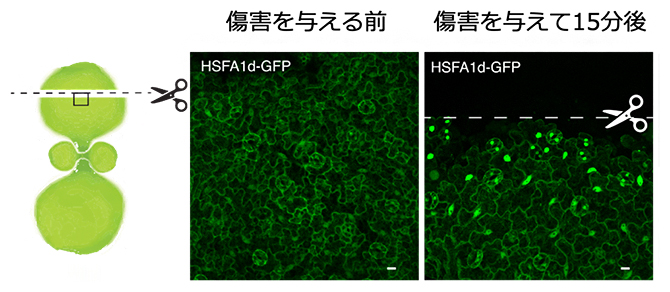
図2 傷害ストレスによって誘導されるHSFA1タンパク質の核内移行
シロイヌナズナの子葉において傷害ストレスを与えた位置(点線)とHSFA1d-GFP融合タンパク質の細胞内局在観察を行った位置(黒枠)を示すモデル図(左)。傷害前(中央)には細胞質に局在していたHSFA1 d -GFP融合タンパク質が傷害後(右)15分以内に核内へ蓄積する様子を示す。写真は子葉表皮細胞の共焦点顕微鏡像。緑色蛍光はHSFA1 d -GFP融合タンパク質の細胞内局在を示す。スケールバー:10マイクロメートル(μm、1μmは100万分の1メートル)。
HSFA1の機能を詳しく調べるために、HSFA1の機能を欠損させたhsfa1多重変異体[10]と活性を亢進(こうしん)させたHSFA1過剰発現体[11]を用いて、傷害後の再生能力への影響を評価しました。その結果、HSFA1の機能が失われると、傷ついた組織からのカルス形成[12]や茎葉再生[13]が著しく低下する一方で、HSFA1の活性が高まると再生能力が顕著に向上することが明らかになりました(図3)。
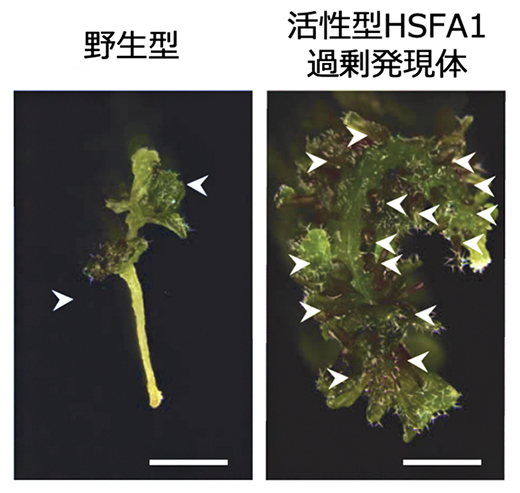
図3 HSFA1の活性化による茎葉再生能力の向上
植物ホルモン添加培地上で培養すると、傷害を受けた胚軸(はいじく)組織片から茎葉が再生する。野生型では通常2個程度の茎葉(白矢印)が形成されるのに対し、活性型HSFA1を過剰発現させた組織片では茎葉再生能力が著しく向上する。スケールバー:2mm。
さらに、HSFA1に依存して発現が変化する遺伝子を探索するため、野生型とhsfa1多重変異体を用いてRNA-seq解析を行いました。その結果、傷害直後から24時間経過する間にHSFA1依存的に誘導される遺伝子群が同定され、その中には細胞リプログラミングに関与する既知の因子が多数含まれていることが分かりました(図4)。
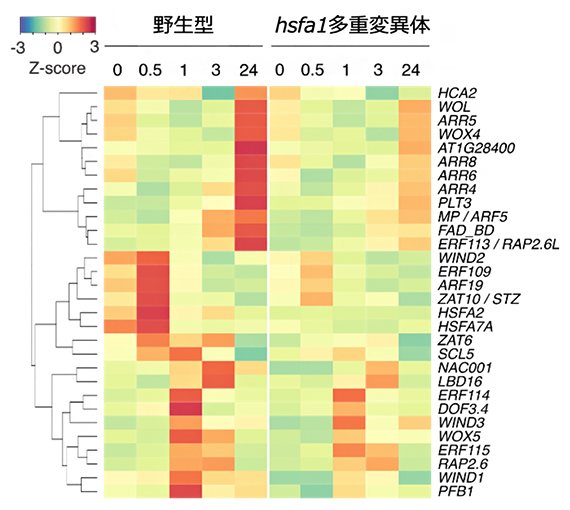
図4 傷害後にHSFA1依存的に発現誘導される遺伝子群
傷害直後から24時間までに野生型で発現誘導される遺伝子のうち、hsfa1多重変異体では発現上昇が抑制される遺伝子の例を示す。HSFA1の下流因子として知られるHSFA2、HSFA7A遺伝子に加え、WIND1、PLT3などの細胞リプログラミング関連遺伝子が含まれる。Z-scoreは赤いほど遺伝子発現が上昇し、青いほど減少することを示す。
これらの遺伝子がHSFA1によって直接制御されているかを検証した結果、HSFA1はWIND1、PLT3などのリプログラミング関連遺伝子のプロモーター領域[14]に直接結合し、それらの発現を誘導することが明らかとなりました。さらに、新たにZAT6遺伝子がリプログラミングに関与すること、HSFA1がその発現も制御することが分かりました。これらの結果から、HSFA1が傷害シグナルを受けて再生に必要な転写ネットワークを迅速に立ち上げることが示されました。
また、HSFA1の活性制御機構についても解析を行った結果、HSFA1の転写活性は、SUMO化[15]による制御を受けて抑制されることが示されました。これにより、HSFA1の活性が過剰にならないよう精緻(せいち)に制御されている可能性が示され、傷害応答における転写制御の新たな調節機構が明らかになりました。
以上の結果から、HSFA1は熱ストレス応答因子にとどまらず、傷害シグナルを細胞リプログラミングへと変換する中核因子として機能することが示されました。
今後の期待
本研究により、傷害という外部刺激がどのようにして細胞リプログラミングを引き起こし、植物の再生につながるのか、その分子基盤の一端が明らかになりました。特に、HSFA1が再生プログラムの上流で機能する転写ハブとして働くことを示した点は、植物再生制御に関する理解を大きく前進させるものです。今後は、HSFA1を介した転写ネットワークの詳細な制御機構や、他のストレス応答経路との相互作用を解明することで、植物がさまざまな条件下で再生能力を発揮する仕組みの全体像が明らかになると期待されます。
また、本研究で得られた知見は、穀物や野菜、果樹などにおける再生・育種技術の高度化や、組織培養・遺伝子導入技術の改善など、応用面への展開が期待されます。特に再生が困難な植物種においてリプログラミング能力を高める技術の開発に貢献することが期待されます。
科学技術振興機構(JST)先端国際共同研究推進事業(ASPIRE)および革新的GX技術創出事業(GteX)の支援を受け、ゲント大学・VIB植物システムズバイオロジーセンター(ベルギー)との国際共同研究を行い、本成果につながりました。
本研究成果は、国際連合が定めた17の目標「持続可能な開発目標(SDGs)[16]」のうち、「2. 飢餓をゼロに」および「15. 陸の豊かさも守ろう」に貢献するものです。
補足説明
- 1.細胞リプログラミング
分化した細胞が性質を変化させ、別の細胞状態へと転換する現象。植物細胞は傷害などの刺激により、再び増殖を開始したり、新たな分化運命を獲得したりして、新たな組織や器官を形成する能力を持つ場合が多い。 - 2.熱ストレス応答
高温環境にさらされた際に生じる生体反応。熱ショックタンパク質などの発現が誘導され、タンパク質の変性を防ぐなど細胞を保護する仕組みが働く。 - 3.HSFA1
熱ストレス応答を制御する代表的な転写因子群。平常時には細胞質に局在し、熱ストレスに応答して核内に移行し、多数の標的遺伝子の発現を活性化するマスター制御因子として機能する。シロイヌナズナにはHSF A1a、HSF A1b、HSF A1d、HSF A1eの四つのホモログ(相同体)が存在し、協調的に下流のHSFAやHSFBなどの他のHSFホモログや、細胞を保護する役割を持つ熱ショックタンパク質HEAT SHOCK PROTEIN (HSP)などの遺伝子発現を制御する。HSFA1はHEAT SHOCK FACTOR A1の略。 - 4.WIND1
傷害ストレスによって誘導され、細胞リプログラミングを促進することで、カルス形成([11]参照)や茎葉再生([12]参照)に重要な役割を果たす転写因子。WIND1はWOUND INDUCED DEDIFFERENTIATION1の略。 - 5.PLT3
根の発生制御などに重要で、細胞の運命決定や組織形成に関与する転写因子。傷害ストレスによっても誘導され、細胞リプログラミングの促進に重要な役割を果たす。PLT3はPLETHORA3の略。 - 6.ZAT6
ストレス応答に関与することが知られており、本研究では新たに細胞リプログラミングへの関与が示された転写因子。ZAT6はZINC FINGER OF ARABIDOPSIS THALIANA6の略。 - 7.転写ネットワーク
複数の転写因子と標的遺伝子が相互に関係しながら遺伝子発現を制御する仕組み。外部刺激に応じて遺伝子発現を統合的に変化させ、細胞の状態や機能を制御する。 - 8.ホモログ(相同体)
共通の祖先に由来し、塩基配列やアミノ酸配列が類似している遺伝子やタンパク質の総称。多くの場合、似た機能を担う。 - 9.RNA次世代シーケンス(RNA-seq)
次世代シーケンサーを用いて、細胞内で発現しているRNAの種類と量を網羅的に解析する手法。RNA-seqはRNA sequencingの略。 - 10.hsfa1多重変異体
HSFA1遺伝子群の複数の機能を欠損させた変異体。今回の研究では主に、ゲノム編集によってHSF A1a、HSF A1b、HSF A1dを欠損させた3重変異体を用いて解析を行った。 - 11.活性を亢進(こうしん)させたHSFA1過剰発現体
抑制ドメイン(領域)を欠損させた活性型HSFA1を、通常よりも高いレベルで発現させた植物体。HSFA1の機能を強めた状態での表現型を解析するために用いられる。 - 12.カルス形成
植物組織から細胞塊が形成される過程。傷害部位で自然に形成されるほか、組織培養では植物ホルモンの作用により人工的に誘導することができる。再生の初期段階として重要であり、新たな器官形成の出発点となる。 - 13.茎葉再生
植物組織から新たに茎や葉が形成される現象。組織培養などにより誘導され、植物の再生能力を評価する指標として用いられる。近年の研究から、この過程には傷害ストレスが重要であることが明らかになっている。 - 14.プロモーター領域
遺伝子発現を制御するDNA配列の一部で、遺伝子の上流に存在することが多い。転写因子が結合することで、遺伝子の発現のオン・オフや強弱が調節される。 - 15.SUMO化
タンパク質に、SUMOと呼ばれる小型タンパク質が共有結合的に付加される翻訳後修飾の一種。タンパク質の活性や安定性、細胞内局在を調節し、ストレス応答や発生制御に関与する。SUMOはSmall Ubiquitin-like Modifierの略。 - 16.持続可能な開発目標(SDGs)
2015年9月の国連サミットで採択された「持続可能な開発のための2030アジェンダ」にて記載された2016年から2030年までの国際目標。持続可能な世界を実現するための17の目標、169のターゲットから構成され、発展途上国のみならず、先進国自身が取り組むユニバーサル(普遍的)なものであり、日本としても積極的に取り組んでいる(外務省のホームページから一部改変して転載)。
国際共同研究グループ
理化学研究所 環境資源科学研究センター 細胞機能研究チーム
特別研究員(研究当時、現 客員研究員)
ダンカン・コールマン(Duncan Coleman)
(現 ゲント大学VIB植物システムズバイオロジーセンター(ベルギー)研究員)
上級研究員 岩瀬 哲(イワセ・アキラ)
(科学技術振興機構(JST)さきがけ研究員)
テクニカルスタッフⅠ 河村 彩子(カワムラ・アヤコ)
テクニカルスタッフⅡ 竹林 有理佳(タケバヤシ・アリカ)
特別研究員(研究当時)陳 渝(チン・ユウ)
訪問研究員 マックス・ミンネ(Max Minne)
訪問研究員(研究当時)デイビッド・ファベロ(David S. Favero)
研修生(研究当時)高橋 達也(タカハシ・タツヤ)
特別研究員(研究当時)池内 桃子(イケウチ・モモコ)
(現 奈良先端科学技術大学院大学 バイオサイエンス領域 教授)
チームディレクター 杉本 慶子(スギモト・ケイコ)
(東京大学 大学院理学系研究科 教授)
宇都宮大学 バイサイエンス教育研究センター
教授 児玉 豊(コダマ・ユタカ)
中部大学 応用生物学部
教授 鈴木 孝征(スズキ・タカマサ)
東京大学 大学院農学研究科
教授 篠崎(山口)和子(シノザキ(ヤマグチ)・カズコ)
大学院生(研究当時)大濱 直彦(オオハマ・ナオヒコ)
ライプニッツ野菜・鑑賞植物研究所(ドイツ)
教授 フィリップ・ウィグ(Philip A Wigge)
研究員 カティア・ジャガー(Katja E Jager)
研究員 マオリン・ペン(Maolin Peng)
ゲント大学VIB植物システムズバイオロジーセンター(ベルギー)
教授 リーヴェン・デ・ヴェルデル(Lieven De Veylder)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費助成事業若手研究「Molecular analysis of novel wound-induced regulators of shoot regeneration in Arabidopsis(研究代表者:ダンカン・コールマン、JP22K15150)」、同新学術領域研究(研究領域提案型)「傷害ストレス誘導性カルスの幹細胞新生メカニズム(研究代表者:岩瀬哲、JP20H04893)」、同学術変革領域研究(B)「プラスチド相転換を制御する因子の機能解析(研究代表者:岩瀬哲、JP22H05075)」、同学術変革領域研究(A)「不均一環境変動に対する植物のレジリエンスを支える多層的情報統御の分子機構(研究代表者:松下智直、研究分担者:児玉豊、JP20H05905)」「植物の環境レジリエンスを支える傷害修復機構(研究代表者:杉本慶子、JP20H05911)」、同特別研究員奨励費「植物メリステムで細胞増殖をエピジェネティックに制御する仕組みの解明(研究代表者:杉本慶子、JP23KF0089)」、同基盤研究(B)「植物の器官再生を支える細胞リプログラミング制御機構の解明(研究代表者:杉本慶子、JP24K02051)」、科学技術振興機構(JST)戦略的創造研究推進事業さきがけ「植物分子の機能と制御」研究領域「低分子化合物から読み解く植物細胞の分化全能性(研究代表者:岩瀬哲、JPMJPR20D2)」、同先端国際共同研究推進事業(ASPIRE)「植物の誘導リプログラミングに立脚した新規バイオエコノミー基盤の創出(研究代表者:杉本慶子、JPMJAP2306)」、同革新的GX技術創出事業(GteX)バイオものづくり領域「先端的植物バイオものづくりの基盤の構築(研究代表者:大熊盛也、主たる共同研究者:杉本慶子、JPMJGX23B0)」の助成を受け、文部科学省国費外国人留学生制度(採用対象者:ダンカン・コールマン)(研究当時)、JSPS外国人特別研究員制度(採用対象者:マックス・ミネ)による支援を受けて行われました。
原論文情報
- Duncan Coleman, Akira Iwase, Ayako Kawamura, Arika Takebayashi, Yu Chen, Max Minne, Katja E. Jager, Maolin Peng, Yutaka Kodama, David S. Favero, Tatsuya Takahashi, Momoko Ikeuchi, Takamasa Suzuki, Naohiko Ohama, Kazuko Yamaguchi-Shinozaki, Philip A. Wigge, Lieven de Veylder, Keiko Sugimoto, "Wounding activates the HSFA1 transcription factors to promote cellular reprogramming in Arabidopsis", The Plant Cell, 10.1093/plcell/koag124
発表者
理化学研究所
環境資源科学研究センター 細胞機能研究チーム
チームディレクター 杉本慶子(スギモト・ケイコ)
特別研究員(研究当時、現 客員研究員)ダンカン・コールマン(Duncan Coleman)
 杉本 慶子、岩瀬 哲、河村 彩子、マックス・ミンネ
杉本 慶子、岩瀬 哲、河村 彩子、マックス・ミンネ
JST事業に関する問い合わせ
科学技術振興機構 国際部 先端国際共同研究推進室
荒川 敦史(アラカワ・アツシ)
Tel: 03-6261-1994
Email: aspire@jst.go.jp
報道担当
理化学研究所 広報部 報道担当
お問い合わせフォーム
科学技術振興機構 広報課
Tel: 03-5214-8404
Email: jstkoho@jst.go.jp