理化学研究所(理研)生命医科学研究センター 免疫転写制御研究チームの奥山 一生 上級研究員(研究当時)、谷内 一郎 チームディレクターらの共同研究グループは、ゲノム上の非メチル化CpG島[1]と呼ばれる領域に結合するタンパク質Cxxc1が、免疫応答を誘導するT細胞[2]の細胞特異的[3]転写因子[4]であるタンパク質Bcl11bと相互作用することで、リンパ球の一種であるT細胞に特異的な遺伝子群の発現を制御する仕組みを明らかにしました。
本研究成果は、Cxxc1が細胞特異的な遺伝子群の発現をどのように制御するのかという理解を大きく前進させるものと期待されます。
ゲノム上にはCpG島と呼ばれる特徴的な領域が存在します。このCpG島はメチル化[1]された領域とほとんどされていない領域(非メチル化CpG島)に分類されます。非メチル化CpG島は遺伝子の発現を制御するプロモーター[5]に広く観察され、特に、細胞の生存や増殖などに不可欠なハウスキーピング遺伝子[6]のプロモーターに広く分布しています。Cxxc1は非メチル化CpG島に結合することで、これらの遺伝子の発現を制御することが知られていました。今回、共同研究グループは、T細胞の分化過程において、胸腺内のT前駆細胞[7]ではCxxc1が、非メチル化CpG島ではなく、プロモーター上の標的DNA配列に結合したBcl11bを介してプロモーターに結合し、T細胞に特異的な遺伝子の発現を制御していることを発見しました。
本研究成果は、科学雑誌『Genes & Development』8月1日号掲載に先立ちオンライン版(6月25日付)に掲載されました。
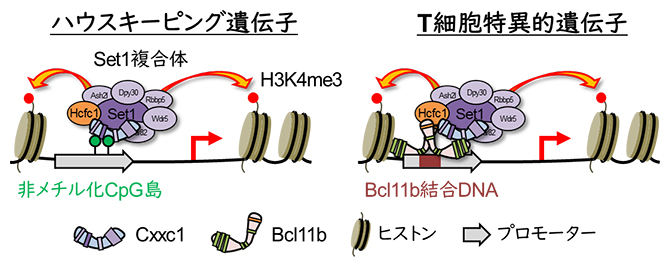
Cxxc1の従来知られていた遺伝子発現の制御機構(左)と今回明らかになった新規の制御機構
背景
細胞の分化・発生過程では、発生段階に応じてクロマチン構造[8]が大きく変化します。クロマチン構造の開閉は遺伝子発現の制御に直結しており、必要な遺伝子領域を開き、不要な領域を閉じることが、正しい遺伝子発現プログラムの確立に不可欠です。T細胞は胸腺で分化(T細胞分化[9])し、骨髄由来の造血幹細胞から生じるDN細胞[9]が、DP細胞[9]を経て、成熟した2種類のSP細胞[9]へと発生段階を進みます。DN細胞からDP細胞への移行では、クロマチン構造が大きく再構成され、それに伴って多くの遺伝子の発現も切り替わることが知られています。
クロマチン構造の形成は、転写因子、クロマチン再構成因子、ヒストン修飾因子などの協調作用によって制御されます。クロマチン再構成因子やヒストン修飾因子は多くの細胞で共通して利用される一方、転写因子はしばしば細胞特異的に発現します。Bcl11bはT細胞に特異的な転写因子であり、2017年に谷内チームディレクターらは、Bcl11bのカルボキシ(C)末端に位置する亜鉛フィンガー(ZnF)構造[10]を欠損させると、DP細胞におけるクロマチン構造の形成が破綻することを報告しました注1)。これらの知見は、Bcl11bのC末端ZnF構造が、クロマチン再構成因子やヒストン修飾因子を結合・動員する上で重要な役割を担っていることを示唆しています。
- 注1)2017年9月26日プレスリリース「T細胞分化を管理する転写制御機構を解明」
研究手法と成果
Bcl11bのC末端ZnF構造がどのようなタンパク質と結合するのかを明らかにするため、共同研究グループは質量分析[11]を用いた網羅的解析を行いました。正常なBcl11bと、C末端ZnFを欠損したBcl11bに結合するタンパク質を精製、同定し、比較したところ、C末端ZnFを欠損したBcl11bではCxxc1との結合能力が大きく低下していることが分かりました。
Cxxc1は非メチル化CpG島に結合し、ヒストン修飾酵素であるSet1複合体を動員してヒストンのメチル化を誘導します。また、DNAをメチル化する酵素タンパク質Dnmt1と結合することでDNAメチル化にも関与します。Cxxc1はT細胞分化に必須であり、特にSet1複合体の動員が重要であることが報告されています注2、3)。
T細胞分化におけるCxxc1の役割を調べるため、共同研究グループはT細胞特異的にCxxc1遺伝子を欠損させたマウスを作製しました。その解析から、Cxxc1欠損マウスの胸腺ではDN細胞からDP細胞への分化が著しく障害されていることが明らかになりました。さらに、Cxxc1欠損DP細胞のクロマチン構造を解析したところ、正常なDP細胞で形成されるべきクロマチン構造が構築されず、DN細胞に類似したクロマチン状態が維持されていることが判明しました。このクロマチン構造の破綻は遺伝子発現にも反映されており、本来DP細胞で高発現する遺伝子の発現が低下し、逆にDN細胞に特異的な遺伝子の発現がDP細胞で持続していました。これらの結果は、Cxxc1がDP細胞に特異的なクロマチン構造と遺伝子発現プログラムの確立に不可欠であることを示しています。
次に、その分子メカニズムを明らかにするため、Cxxc1およびBcl11bのゲノム上の結合領域を解析しました。その結果、Cxxc1が結合する領域の約70%にはBcl11bが同時に存在していることが分かりました。さらに、Bcl11b欠損細胞でCxxc1の結合を評価したところ、本来Cxxc1が結合していた領域の約40%からCxxc1が離脱していました。これは、これらの領域へのCxxc1の結合がBcl11bに依存していることを示しています。このBcl11bに依存的な領域にはT細胞関連遺伝子のプロモーターが多く含まれ、CpG島を持たない領域であることが特徴でした。一方、Bcl11bが存在しなくてもCxxc1の結合が維持された領域には、代謝やタンパク質翻訳に関わる遺伝子群のプロモーターが多く、CpG島が広く分布していました。これらの結果から、Cxxc1の標的領域への結合には「CpG島を介した結合」と「Bcl11bに依存した結合」の2種類が存在し、前者はハウスキーピング遺伝子、後者はT細胞に特異的な遺伝子のプロモーター領域に対応することが示唆されました(図1)。
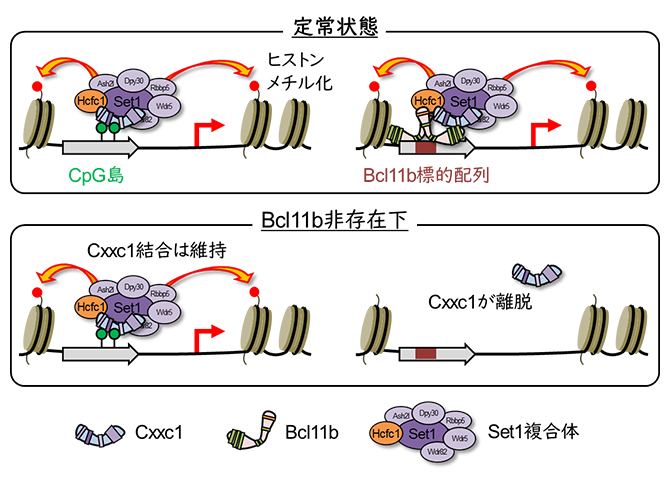
図1 Cxxc1のCpG島およびBcl11b依存的DNA結合
Cxxc1はハウスキーピング遺伝子およびT細胞特異的遺伝子のプロモーター領域に結合する際、それぞれCpG島とBcl11bのC末端ZnFを介すると考えられる。
さらに、B細胞[12]ではB細胞に特異的な転写因子Bcl11aが発現します。Bcl11aはBcl11bと同じBcl11遺伝子ファミリーに属し、C末端ZnF構造を含め高次に類似したタンパク質構造を持ちます。脾臓(ひぞう)B細胞におけるCxxc1とBcl11aのDNA結合領域を解析したところ、Cxxc1結合領域の80%以上にBcl11aが同時に存在していることが明らかになりました。このことから、Cxxc1を介した細胞特異的な遺伝子発現の制御機構はT細胞だけでなく、B細胞の分化過程においても利用されていると考えられます。
- 注2)M. Kiuchi, A. Onodera, K. Kokubo, T. Ichikawa, Y. Morimoto, E. Kawakami, N. Takayama, K. Eto, H. Koseki, K. Hirahara, T. Nakayama, [The Cxxc1 subunit of the Trithorax complex directs epigenetic licensing of CD4+ T cell differentiation], Journal of Experimental Medicine, 2021, DOI: 10.1084/jem.20201690
- 注3)W. Cao, J. Guo, X. Wen, L. Miao, F. Lin, G. Xu, R. Ma, S. Yin, Z. Hui, T. Chen, S. Guo, W. Chen, Y. Huang, Y. Liu, J. Wang, L. Wei, L. Wang, [CXXC finger protein 1 is critical for T-cell intrathymic development through regulating H3K4 trimethylation], Nature Communications, 2016, DOI: 10.1038/ncomms11687
今後の期待
本研究により、Cxxc1がBcl11bと相互作用することでT細胞に特異的な遺伝子の発現制御に寄与することが明らかになりました。また、B細胞の分化過程においても、Bcl11aとCxxc1が協調してB細胞に特異的な遺伝子の発現を調節する可能性が示されました。さらに近年、制御性T細胞に特異的な転写因子Foxp3がCxxc1と相互作用することも報告されています。これらの知見を総合すると、Cxxc1はT細胞やB細胞にとどまらず、さまざまな細胞種において、その細胞特異的な遺伝子発現プログラムの制御に広く関与していることが予測されます。今後、Cxxc1を介した遺伝子発現制御の仕組みをさらに解明することで、細胞の分化異常や発がんなどの疾患の理解につながることが期待されます。
補足説明
- 1.非メチル化CpG島、メチル化
メチル化とは、ゲノム上でシトシン(C)にメチル基(-CH3)が結合するDNA修飾のこと。CpG島とは、ゲノム上でCとグアニン(G)が高密度に存在する領域を指す。特に、ほとんどメチル化されていない領域を、非メチル化CpG島と呼ぶ。 - 2.T細胞
免疫応答を誘導するリンパ球の一群。他の免疫細胞を活性化したり、ウイルス感染した細胞を殺したりする機能を持つ。 - 3.細胞特異的
特定の細胞種(細胞系譜)にのみ、あるいは限られた系譜にのみ限定されるということ。 - 4.転写因子
DNA上の特定の配列に結合し、標的遺伝子の発現を調節するタンパク質の総称。 - 5.プロモーター
遺伝子の「スイッチ」として機能するDNA領域。転写が始まる位置(転写開始点)の直上流に位置し、転写因子や転写制御因子が結合することで遺伝子発現が制御される。 - 6.ハウスキーピング遺伝子
生存・増殖・代謝など、全ての細胞に共通する基本的な機能を担うタンパク質をコードする遺伝子群を指す。細胞特異的な遺伝子とは異なり、ほぼ全ての細胞で恒常的に発現している。 - 7.T前駆細胞
特定の細胞系譜へ分化する能力を持つが、幹細胞のような自己をコピーする能力(自己複製能)は持たない細胞。骨髄の造血幹細胞に由来するT前駆細胞が胸腺へ移動し、胸腺内の環境によりT細胞分化([9]参照)が進む。 - 8.クロマチン構造
ヒストン(八量体タンパク質)にDNAが巻き付いたヌクレオソームを基本単位とし、これがさらに折り畳まれた高次構造。クロマチンは「開いた構造」と「閉じた構造」に大別され、前者では遺伝子発現が活性化され、後者では抑制される。「開いた構造」とはDNAがヒストンから解け、ヌクレオソーム同士の間隔が広がった構造であり、「閉じた構造」とはヌクレオソームが高密度に集積している構造である。 - 9.T細胞分化、DN細胞、DP細胞、SP細胞
T細胞の分化は胸腺内の微小環境によって支持される。細胞の分化段階は、細胞表面に発現するタンパク質を、フローサイトメーターと呼ばれる細胞の大きさや内部構造、細胞膜表面に発現するタンパク質の種類と量を測定できる装置で解析することで同定される。T細胞は、CD4とCD8と呼ばれる二つのタンパク質の発現状態により、両方を発現していないDN細胞(最も未熟)、両方を発現するDP細胞(中間)、CD4あるいはCD8のいずれかのみを発現するCD4 SPおよびCD8 SP細胞(最も成熟)に分類される。 - 10.亜鉛フィンガー(ZnF)構造
亜鉛イオンにより安定化されるタンパク質の特徴的な立体構造の一つ。多くの転写因子に存在し、DNAや他のタンパク質との結合に重要な役割を果たす。 - 11.質量分析
分子の質量を高精度に測定する分析手法で、タンパク質の同定に広く用いられる。 - 12.B細胞
リンパ球と呼ばれる免疫細胞の一種で、抗体(免疫グロブリン)というタンパク質の産生を担う。この抗体がウイルスや細菌、毒素といった異物に特異的に結合して排除する。
共同研究グループ
理化学研究所 生命医科学研究センター
免疫転写制御研究チーム
チームディレクター 谷内 一郎(タニウチ・イチロウ)
上級研究員(研究当時)奥山 一生(オクヤマ・カズキ)
(現 国立がん研究センター研究所 がんRNA研究分野 主任研究員)
研究員(研究当時)ウーセオク・セオ(Wooseok Seo)
テクニカルスタッフⅠ 室井 佐和子(ムロイ・サワコ)
研修生(研究当時)ニン・ホウ(Ning Hou)
融合領域リーダー育成プログラム
上級研究員(研究当時)イボ・ウー(Yibo Wu)
(現 粘膜免疫研究チーム 客員研究員)
免疫器官形成研究チーム
チームディレクター 古関 明彦(コセキ・ハルヒコ)
研究員 伊藤 伸介(イトウ・シンスケ)
愛媛大学 プロテオサイエンスセンター 無細胞生命科学部門
教授 澤崎 達也(サワサキ・タツヤ)
准教授 高橋 宏隆(タカハシ・ヒロタカ)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費助成事業基盤研究(C)「BCL11B変異によるヒト原発性免疫不全症発症機序の解明(研究代表者:奥山一生、20K07459)」、同新学術領域研究(研究領域提案型)「ネオ・セルフ抗原認識をT細胞分化制御に転換する分子機構の解明(研究代表者:谷内一郎、19H04820)」による助成を受けて行われました。
原論文情報
- Kazuki Okuyama, Wooseok Seo, Hirotaka Takahashi, Sawako Muroi, Ning Hou, Shinsuke Ito, Tatsuya Sawasaki, Haruhiko Koseki, Yibo Wu, and Ichiro Taniuchi, "Bcl11-Cxxc1 axis regulates stage-specific chromatin accessibility during lymphocyte development", Genes & Development, 10.1101/gad.353082.125
発表者
理化学研究所
生命医科学研究センター 免疫転写制御研究チーム
上級研究員(研究当時)奥山 一生(オクヤマ・カズキ)
チームディレクター 谷内 一郎(タニウチ・イチロウ)
 奥山 一生
奥山 一生
 谷内 一郎
谷内 一郎
報道担当
理化学研究所 広報部 報道担当
お問い合わせフォーム