理化学研究所(理研)環境資源科学研究センター 植物ゲノム発現研究チームの関 原明 チームディレクター、上田 実 研究員、ミュンスター大学のイリス・フィンケマイアー 教授、フロリアン・コーニック 大学院生らの国際共同研究グループは、植物の耐塩性に関わるヒストンアセチル化[1]サイト(位置)を同定しました。
本研究成果は、植物の塩ストレス耐性を高めるための品種改良や化合物などを介した環境ストレス応答を操作する方法の開発への貢献が期待されます。
今回、国際共同研究グループは、モデル植物であるシロイヌナズナにおいて、ヒストンのアセチル化レベルの調節に関わり、耐塩性を高めることが知られていたヒストン脱アセチル化酵素(HDAC)[2]の変異体、hda19変異体のアセチローム解析[3]を行いました。その結果、hda19変異体では、DNAを細胞内に整然と折り畳む働きがあるタンパク質の一つである、ヒストンH3.3バリアントの27番目と36番目の二つのリシン残基のアセチル化(H3.3K27acK36ac)が強く誘導されていることを発見しました。アセチル化リシン残基と電荷的な性質と構造が類似しているグルタミン残基に置換したアセチル化ミミック(疑似体)を植物に発現させることでも、耐塩性を付与できました。この結果は、この高アセチル化が、耐塩性を規定するヒストンコード[4]であることを示唆します。hda19が高アセチル化を介して耐塩性を示すために必須な遺伝子も同定しました。
本研究は、従来の耐塩性関係のパターンと異なるアセチル化修飾が耐塩性獲得の鍵になることを示しており、耐塩性に関わるエピジェネティック制御[5]機構の理解を大きく前進させるものです。
本研究は、科学雑誌『Proceedings of the National Academy of Sciences of the United States of America(PNAS)』オンライン版(7月13日付)に掲載されました。
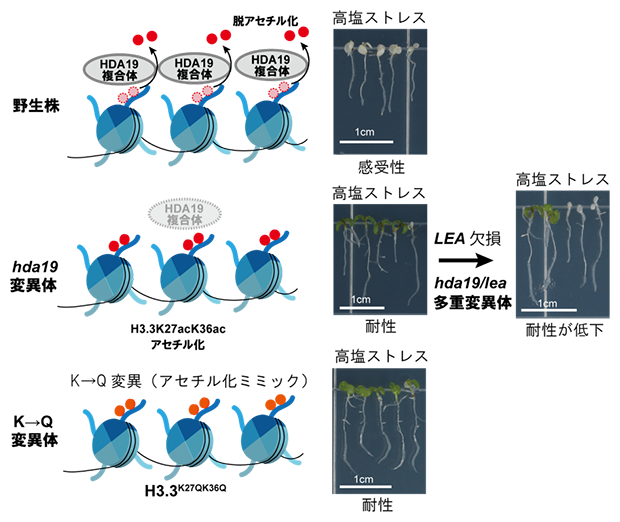
HDA19が調節するアセチル化サイトとhda19変異体が耐塩性になるための必須遺伝子を同定
背景
近年、気候変動は農業生産へ負の影響を与えています。塩害は、台風などの強風によって海水の影響を受ける海岸部だけではなく、岩塩地層を持つ地域や過度の灌漑(かんがい)や乾燥などにより、表層土壌に塩類が集積した地域で発生しており、塩害は干ばつと同様に年々深刻化しています。そのため、植物の耐塩性を向上させる技術開発が世界各国で進められています。
関チームディレクターらは、これまでにエピジェネティック制御因子の一つである、ヒストンアセチル化修飾を調節するヒストン脱アセチル化酵素(HDAC)の制御による耐塩性強化法の開発を進めてきました。モデル植物であるシロイヌナズナのゲノムでは、HDACとして働く遺伝子が多数存在し、遺伝子ファミリー[6]を形成していることが知られています。関チームディレクターらは、HDAC遺伝子ファミリー中で、HDA19の活性を阻害・欠損することで耐塩性が高まることを突き止めていました注1、2)。ただし、四つのコアヒストン上に20以上あるアセチル化修飾を受けるリシン残基のうち、どのリシン残基でのアセチル化修飾をHDA19が調節しているのか不明のままでした。また、ヒストン以外のタンパク質もアセチル化修飾を受けることが知られており、HDA19制御下にある、耐塩性に関わるアセチル化タンパク質の同定が期待されていました。
そこで国際共同研究グループは、hda19変異体においてアセチル化レベルが上昇しているタンパク質の網羅的な解析を進め、その中から耐塩性の鍵となるアセチル化サイトとタンパク質の同定を目指しました。
- 注1)2015年12月24日プレスリリース「植物の耐塩性を高める化合物を発見」
- 注2)2017年11月7日プレスリリース「植物の耐塩性に関わるヒストン脱アセチル化酵素の同定」
研究手法と成果
まず国際共同研究グループは、脱アセチル化酵素として働くHDA19の機能を失っているhda19変異体では、脱アセチル化の機能が弱まることで、一部のタンパク質でアセチル化レベルが高まっていることが予想されるため、植物が子葉に蓄えた栄養に依存した生長から、光合成を行い自立した生長を始めるステージにある発芽誘導後5日目のhda19変異体を材料にアセチローム解析を行いました。その結果、hda19変異体ではヒストンH3.3やLate embryogenesis abundant(LEA)タンパク質など多数のタンパク質においてアセチル化レベルが高まっていることが分かりました。
アセチローム解析から、コアヒストンの中では、ヒストンH3.3バリアントの27番目と36番目の二つのリシン残基(H3.3K27acK36ac)の高アセチル化が検出できたため、H3.3K27acK36acに着目して解析を進めました。シロイヌナズナゲノムは、ヒストンH3を15個の遺伝子にコードしており、タンパク質を構成するアミノ酸配列の違いから、シロイヌナズナでは、H3.1、H3.3、H3.6、H3.7、H3.10、H3.11、CenH3、H3.14、H3.15の各バリアントに分類されています。
数あるH3バリアントの中から、H3.3K27acK36ac高アセチル化が耐塩性に関わることを直接証明するために、アセチル化を受けるリシン残基をグルタミン残基に置換することで、高アセチル化状態を模倣する変異体(アセチル化ミミック)を作出しました。つまり、H3.3の27番目と36番目の二つのリシン残基をグルタミン残基に置換した植物体(H3.3K27QK36Q、K→Q変異体[7])を作出し、そのK→Q変異体の耐塩性を評価しました(図1)。その結果、K→Q変異体はコントロール(対照群)の植物体と比べて、耐塩性を示したことから、H3.3K27acK36ac高アセチル化は耐塩性を高めることが示唆されました。
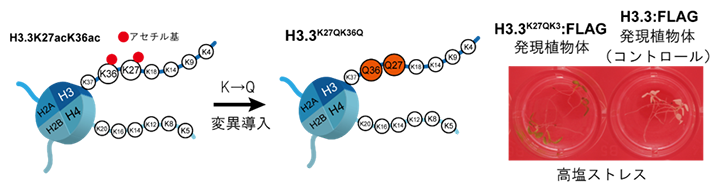
図1 H3.3K27acK36ac高アセチル化ミミックを発現するシロイヌナズナの耐塩性試験
液体培養し,塩ストレスを与えたK→Q変異を導入していないコントロールの植物体は死滅した。一方、K→Q変異を導入したH3.3を発現する植物体は、塩ストレス条件下でも生存可能になった。翻訳産物の検出と後述のChIP-qPCR解析のためFLAGタグをH3.3タンパク質のC末端側に付加している。
さらに、アセチローム解析の結果から、野生株と比べてhda19変異体で高蓄積していることが判明したLEAタンパク質(LEA7、LEA29、RAB18)を欠損したhda19変異体(hda19/lea多重変異体)の耐塩性を調べました。その結果、hda19/lea多重変異体はhda19変異体と比較して、耐塩性が低下していることが分かりました(図2)。つまり、LEA7、LEA29、RAB18はhda19変異体が耐塩性を示すために必須の遺伝子であることも判明しました。
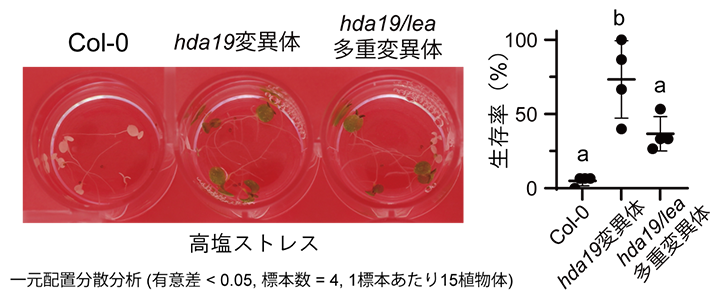
図2 LEA7/LEA29/RAB18を欠損したhda19変異体(hda19/lea多重変異体)の耐塩性低下
液体培養し,塩ストレスを与えた野生株(Col-0)植物体は死滅した。一方、hda19変異体は塩ストレス条件下でも生存可能になった。三つのLEAを欠損させたhda19/lea多重変異体ではhda19変異体と比べて、耐塩性が低下する。右のグラフでは、多重比較(Tukey法)により他の群と比較して有意とされた群については異なるアルファベットをグラフ上に示している。
実際に、H3.3は各LEA遺伝子の転写を調節するためのプロモーター領域に局在していることをクロマチン免疫沈降(ChIP)[8]-定量リアルタイムPCR(qPCR)[9]により確認できました(図3b)。さらに、それぞれの植物体から抽出したRNAをqPCRにより解析した結果、K→Q変異体でのみ各LEA遺伝子の転写(遺伝子発現)が高まっていることが分かりました(図3a)。一般にアセチル化レベルが高まると、遺伝子発現も高まる正の相関を示すことが知られていますが、H3.3K27acK36acアセチル化サイトについてもこの相関がみられ、H3.3K27acK36ac高アセチル化が各LEA遺伝子の転写を高めていることが明らかとなりました。
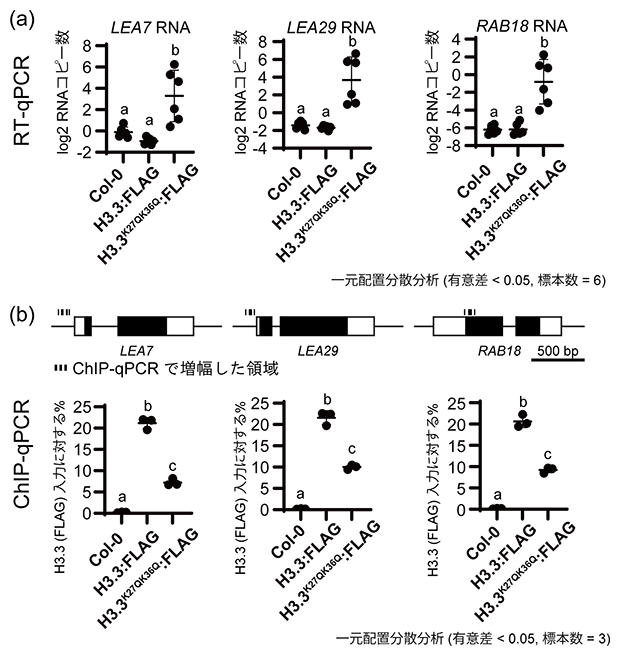
図3 ヒストンH3.3K27acK36ac高アセチル化(K27QK36Q)によるLEA遺伝子群の発現上昇
- (a)各LEA遺伝子の転写産物(RNA)での遺伝子発現定量リアルタイムPCR(RT-qPCR)解析結果。H3.3K27QK36Qを発現する植物体でのみ、各LEA遺伝子の発現量が高まっている。
- (b)遺伝子プロモーターまたは上流配列でのFLAG抗体によるクロマチン免疫沈降-定量リアルタイムPCR(ChIP-qPCR)の解析結果。FLAGタグを融合しているH3.3とH3.3K27QK36Qともに各LEA遺伝子のプロモーター領域に存在している。
各グラフにおいて、多重比較(Tukey法)により他の群と比較して有意とされた群については異なるアルファベットをグラフ上に示している。
今後の期待
今後、H3.3K27acK36acアセチル化制御下にある遺伝子群の機能解析により、化合物などによる緻密な塩ストレス応答操作法の開発や、新規の耐塩性に関わる分子機構の同定が期待されます。
HDACは、特定のアセチル化サイトで脱アセチル化反応を触媒するために、単独で働くわけではなく、特定のアセチル化サイトを認識するために必要なタンパク質であるリーダータンパク質と複合体を形成することで機能していると考えられています。本研究で耐塩性に関わるアセチル化サイト(H3.3K27acK36ac)を同定することができたことから、今後はHDA19と協調してH3.3K27acK36acアセチル化を認識するリーダータンパク質の同定が期待されます。そして、HDA19とそのリーダータンパク質の結合を阻害する化合物を同定することなどで、化合物による塩ストレス応答操作法の開発が期待されます。
今回の研究成果は、国際連合が2016年に定めた17項目の「持続可能な開発目標(SDGs)[10]」のうち「2.飢餓をゼロに」や「13.気候変動に具体的な対策を」などへの貢献が期待できます。
補足説明
- 1.ヒストンアセチル化
ヒストンは、塩基性アミノ酸であるリシンやアルギニン残基が多く含まれていることから、正の電荷を持つ。一方で、DNAは負の電荷を持つことから、ヒストンとDNAは安定した複合体を形成できる。ヒストンアセチル化は、主にリシン残基で起こり、その正の帯電を解消することでDNAの巻きつきを弱めたり、またはタンパク質間相互作用を変化させたりして、転写因子やRNAポリメラーゼがより結合しやすい状態にすることで転写促進に関わると考えられている。ヒストンはこの他にもメチル化やリン酸化など、さまざまな化学修飾を受けることが知られている。 - 2.ヒストン脱アセチル化酵素(HDAC)
アセチル化された部位を加水分解により除去し、ヒストンへのDNAの巻きつきを強めることなどによって転写を抑制する酵素。ヒストン脱アセチル化酵素の機能を抑制するとアセチル化が促進され転写の促進に寄与する。 - 3.アセチローム解析
アセチロームとは、対象とした細胞・器官などに存在している全てのアセチル化タンパク質の集合・総体を意味する。アセチローム解析は、生体内における質量分析装置で検出された各タンパク質のアセチル化レベルを網羅的に調べることを主な目的としている。ヒストン以外のタンパク質もアセチル化修飾を受ける。 - 4.ヒストンコード
ヒストンコードとはヒストンの化学修飾の組み合わせが遺伝子発現調節のための暗号(コード)として機能すること。ヒストンとは染色体を構成する主要なタンパク質である。染色体は、ヒストンH2A、H2B、H3、H4のそれぞれ2分子から成る八量体に、DNAが左巻きに1.65回巻きついた複合体を基本単位(ヌクレオソーム)として存在している。ヌクレオソームからヒストンテールと呼ばれる各ヒストンのアミノ酸末端領域での、メチル化やアセチル化、リン酸化などの化学修飾が、染色体構造に変化をもたらし、遺伝子発現の抑制や活性化を調節する。米国のアリス博士らによりヒストン上の化学修飾のさまざまな組み合わせが暗号(コード)のように働き、各コードが固有の細胞運命を決定するための遺伝子発現調節を担うとするヒストンコード仮説が提唱されている。 - 5.エピジェネティック制御
DNA配列の変化を伴わず、DNAやヒストンへの後天的な化学修飾により制御される遺伝現象。DNAのメチル化や、ヒストンのアセチル化、メチル化などが、後天的な修飾として作用する。 - 6.遺伝子ファミリー
進化の過程で共通の遺伝子から、遺伝子重複などを経てコピー数を増やしたグループ。HDACはさらに、RPD3-like、Sirtuin、HD-tuinの三つのグループ(サブファミリー)に分類されており、HDA19はRPD3-likeに分類される。 - 7.K→Q変異体
リシン残基のアセチル化は、側鎖が持つ正電荷が消失することで分子間、もしくは分子内の相互作用を変化させる。グルタミン残基の側鎖は、長さと形状が異なるものの、アセチル化リシンと電荷が中性である点や、極性アミド基の構造が似ている点があるため、分子生物学や生化学実験では長年「アセチル化ミミック(疑似体)」として利用されている。 - 8.クロマチン免疫沈降(ChIP)
細胞のクロマチンを断片化し、特定の抗体が認識するタンパク質やその化学修飾の状態を含むクロマチン断片を取得する方法。 - 9.定量リアルタイムPCR(qPCR)
測定する試料に含まれる特定の配列を含むDNA量の推定が可能なDNAの増幅法。極めて微量な試料からも高精度に解析できる。 - 10.持続可能な開発目標(SDGs)
2015年9月の国連サミットで採択された「持続可能な開発のための2030アジェンダ」にて記載された2016年から2030年までの国際目標。持続可能な世界を実現するための17のゴール、169のターゲットから構成され、発展途上国のみならず、先進国自身が取り組むユニバーサル(普遍的)なものであり、日本としても積極的に取り組んでいる(外務省ホームページから一部改変して転載)。
国際共同研究グループ
理化学研究所 環境資源科学研究センター
植物ゲノム発現研究チーム
チームディレクター 関 原明(セキ・モトアキ)
研究員 上田 実(ウエダ・ミノル)
テクニカルスタッフⅠ 高橋 聡史(タカハシ・サトシ)
テクニカルスタッフⅠ 石田 順子(イシダ・ジュンコ)
研修生(研究当時)阿部 貴裕(アベ・タカヒロ)
天然物生合成研究ユニット
ユニットリーダー 高橋 俊二(タカハシ・シュンジ)
研究員 坂井 克行(サカイ・カツユキ)
テクニカルスタッフⅠ 髙木 海(タカギ・ヒロシ)
東京薬科大学 生命科学部 細胞情報科学研究室
教授 伊藤 昭博(イトウ・アキヒロ)
ミュンスター大学(ドイツ)
教授 イリス・フィンケマイアー(Iris Finkemeier)
大学院生 フロリアン・コーニック(Florian Kotnik)
大学院生 ユルゲン・アイリッヒ(Juergen Eirich)
テュービンゲン大学(ドイツ)
教授 ディルク・シュバルツァー(Dirk Schwarzer)
大学院生(研究当時)ユリアン・ザイデル(Julian Seidel)
研究支援
本研究は、理研横断プロジェクト「エピゲノム操作(研究代表者:眞貝洋一)」により実施し、科学技術振興機構(JST)先端国際共同研究推進事業ASPIRE「植物のレジリエンスを強化する国際研究センター(研究代表者:関原明)」、同戦略的創造研究推進事業CREST「エピゲノム制御ネットワークの理解に基づく環境ストレス適応力強化および有用バイオマス産生(研究代表者:関原明)」、同革新的GX技術創出事業GteX「先端的植物バイオものづくり基盤の構築(研究代表者:大熊盛也)」、日本学術振興会(JSPS)科学研究費助成事業基盤研究(C)「HDACの遺伝的・化学的改変による環境ストレス応答最適化法の開発(研究代表者:上田実)」による助成を受けて行われました。
原論文情報
- Florian Kotnik*, Minoru Ueda*, Akihiro Ito, Junko Ishida, Satoshi Takahashi, Katsuyuki Sakai, Hiroshi Takagi, Julian Seidel, Takahiro Abe, Juergen Eirich, Shunji Takahashi, Dirk Schwarzer, Motoaki Seki#, Iris Finkemeier #(*:共同筆頭著者、#:共同責任著者), "HDA19-mediated deacetylation of histone H3.3 lysine 27 and 36 regulates plant sensitivity to salt stress", Proceedings of the National Academy of Sciences of the United States of America(PNAS), 10.1073/pnas.2534315123
発表者
理化学研究所
環境資源科学研究センター 植物ゲノム発現研究チーム
チームディレクター 関 原明(セキ・モトアキ)
研究員 上田 実(ウエダ・ミノル)
ミュンスター大学(ドイツ)
教授 イリス・フィンケマイアー(Iris Finkemeier)
大学院生 フロリアン・コーニック(Florian Kotnik)
 左から上田 実、関 原明
左から上田 実、関 原明
 左からフロリアン・コーニック、イリス・フィンケマイアー
左からフロリアン・コーニック、イリス・フィンケマイアー
報道担当
理化学研究所 広報部 報道担当
お問い合わせフォーム