理化学研究所(理研)脳神経科学研究センター 認知分散処理研究チームのシュミット・ルーカス・イアン チームディレクター、フォスフォード・パトリック 特別研究員(研究当時)、メイ・ハオ 特別研究員、中島 美保 副チームディレクターらの研究チームは、不確実な状況下で直感を働かせるとき、脳がどのように情報を処理しているのかを解明しました。私たちは、外界から得られる感覚情報が不十分な場合でも過去の経験をもとに、推測(直感)して行動します。本研究はこの判断材料となる「感覚経験」が脳内でどのように保存され、さらに新しい経験によってどのように更新されていくのか、そのプロセスを突き止めました。特に脳内に蓄積された過去の統計的情報が外界の状況変化に応じて少しずつ書き換えられ、新たな情報を取り入れる態勢を整える神経機構を明らかにしました。
本研究成果は、変化の激しい環境でいかに適応していくのかという情報のスクリーニング過程と直結し、将来的には思い込みによる認知バイアスの制御、情報に惑わされない精神的健康の維持、幻聴の治療開発といった幅広い分野への社会実装に貢献すると期待されます。
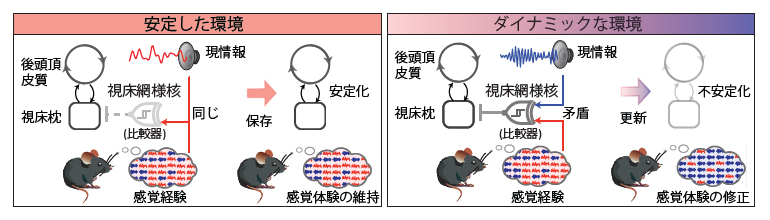
研究チームは、最近聞いた音の傾向が後頭頂皮質[1]―視床枕(ししょうちん)[2]回路に「感覚経験」情報として保存され、曖昧な情報のもとでの推測(直感)の基盤となっていることを発見しました。また、状況の変化とともに「感覚経験」情報を段階的にリセットすることで判断にバイアスがかからないようにするために、脳内では、視床網様核[3]が「感覚経験」情報と現在の情報を照合する比較器として機能し、両者のミスマッチを検知すると、「感覚経験」情報が状況の変化とともに段階的に更新されて、脳がより新たな情報を取り入れるモードに切り替わることを発見しました。
本研究は、科学雑誌『Neuron』オンライン版(5月29日付:日本時間5月30日)に掲載されました。
直感を支える感覚経験の更新メカニズム
背景
曖昧な情報のもとで判断を迫られるとき、誰しも直感で判断することがあります。このような直感は多くの場合、過去の経験、特に最近起きた出来事の蓄積で構築した脳の内的情報に基づいています。内的情報は判断の指針となりますが、過剰に強固になると判断にバイアスがかかり、現実を正しく理解できなくなります。
例えば、ドアベルが鳴ったとき、脳はドアベルの音を「感覚経験」の内的情報として蓄積し、今後ドアベルを聞き逃さないようにします。その一方で、単なる雑音がこの内的情報の影響を受けてドアベルのように聞こえ、正しく判断できないこともあります。このように「感覚経験」の内的情報は現在の感覚処理にも影響を与えます。これが深刻化すると妄想や幻聴といった症状の一因にもなり得るため、内的情報を使いつつも新しい情報に合わせて内的情報を上書きする適応性が重要であり、変化に応じて無意識にでも上書きを指示する動的な調整機能が不可欠です。
これまでの研究により、内的情報が後頭頂皮質に保存されていることが示唆され注1)、内的情報と現在の情報の違いを検知するニューロンの存在は発見されていました注2)。しかし、「脳内にて内的情報が更新される瞬間」を捉えることが極めて困難なため、脳がどのように現在の状況に合わせて、内的情報を保持し、段階的に更新しているのかは不明でした。
今回、研究チームは、独自に開発した内的情報ダイナミック更新パラダイム(行動課題、刺激提示法)に複数の脳領域の同時活動測定・制御法を組み合わせることで、どのように内的情報が状況に応じて更新されるのか、その神経機構、計算原理の解明に挑みました。
- 注1)Athena Akrami et al. Posterior parietal cortex represents sensory history and mediates its effects on behaviour. Nature, 2018 Feb 15.
- 注2)Rajesh P. N. Rao, Dana H. Ballard. Predictive coding in the visual cortex: a functional interpretation of some extra-classical receptive-field effects. Nature Neuroscience, January 1999.
研究手法と成果
研究チームは音刺激判別の行動課題を新たに構築しました。この課題では、マウスに連続して流れる2種類の音(高周波と低周波)から成る音刺激から、どちらの音の割合が大きいかを聞き分けさせ、高周波の割合が多いなら右、低周波の割合が多いなら左という特定の方向へ行動させるようにすることで、脳が感覚情報をどのように知覚したのか調べる課題です。この行動課題では、そのとき提示された音刺激のみに頼って意思決定ができる試行(現情報試行)と、現情報(2種類の音の割合が同じ)だけでは判断ができないので、最近の傾向(「感覚経験」、前の5試行における音刺激の合計で、より多く流れた音が指す方向)を基に意思決定する試行(経験試行)がランダムに現れます(図1A)。そのため、マウスは現情報を適切に処理しつつ、最近の傾向も「感覚経験」として脳内に蓄積する必要があります。
まず、この「感覚経験」が先行研究と同様に後頭頂皮質に保存されているのかを検証するために、この行動課題の試行間に後頭頂皮質の活動を光遺伝学法[4]を用いて抑制すると、この抑制群のマウスの行動に二つの特徴が見られました(図1B)。
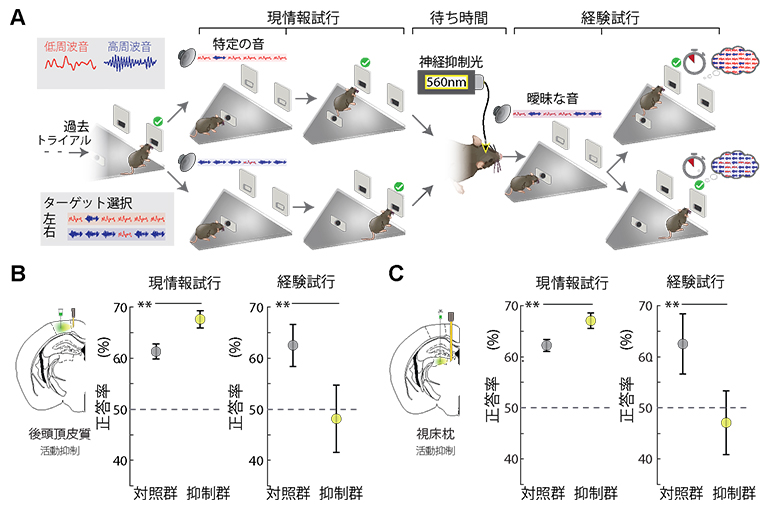
図1 最近経験した感覚情報は視床枕と後頭頂皮質ループ内に保存
- (A) マウスの行動課題。現在流れている音刺激を基に、左右の穴のうち音が示す穴(低周波音が多いときは左、高周波音が多いときは右)に鼻を入れるという意思決定をする現情報試行と、最近の試行(感覚経験)情報を基に判断する経験試行がランダムに現れる。
- (B、C) 視床枕もしくは後頭頂皮質の活動抑制を試行間(待ち時間)で行うと、現情報試行の正答率は上がるが、経験試行の正答率は下がる。「**」有意水準1%での有意差あり。
第一に、経験試行の正答率が下がりました。この結果は、経験試行の遂行に必要な「感覚経験」(直前の5試行合計の中で多く流れた音刺激)情報が、後頭頂皮質の活動抑制によって消滅し、その情報を生かした正しい意思決定ができなかったことを示します。そのため、「感覚経験」情報は後頭頂皮質に保存されていることが分かりました。
第二に、現情報試行の正答率は上がりました。この結果は、マウスが後頭頂皮質に保存された「感覚経験」情報によって現情報処理にバイアスがかかり、正しい判断ができなかった可能性を示しています。
次に、「感覚経験」情報が後頭頂皮質内回路だけで保持されているのか、もしくは、ある程度長期間保持するために他の回路も関わっているのか、検証するために、後頭頂皮質と相互回路を形成する視床枕の活動も光遺伝学法で抑制しました。その結果、後頭頂皮質と同様の結果が得られ、「感覚経験」情報の保持には視床枕も必要なことが分かりました(図1C)。
次に、「感覚経験」情報がどのように後頭頂皮質・視床枕に保存されているのかを明らかにするために、行動課題を遂行中のマウス後頭頂皮質の神経活動を記録しました(図2A)。後頭頂皮質の活動には「感覚経験」情報がシンボル化(表象化)されていますが、この表象化は、視床枕の活動を抑制したことで非常に弱くなり(図2B、C)、「感覚経験」情報が後頭頂皮質・視床枕ループで保持されていることが示唆されました(図2D)。また、この「感覚経験」情報は、行動課題と関係ない受動的に聞く音情報の履歴でも、後頭頂皮質・視床枕に蓄積されることも示したため、無意識に判断する直感に関与している可能性が高いことを示唆しています。
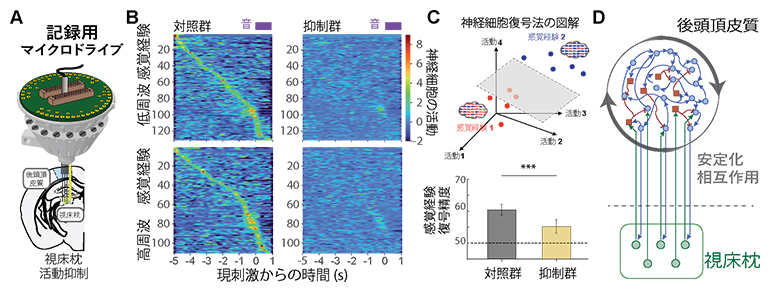
図2 後頭頂皮質に保存された感覚経験情報の視床枕の活動抑制による弱体化
- (A)マイクロドライブ(小型記録装置)を用いた後頭頂皮質の活動測定の模式図。一部の試行間(待ち時間)で視床枕の活動抑制も同時に行った。
- (B)後頭頂皮質のニューロン群にシンボル化(表象化)された「感覚経験」情報は、視床枕の活動抑制によってほぼ消滅した。
- (C)神経細胞の活動から感覚経験の内容を復号化する。(上)異なる「感覚経験」状態(赤は低周波数が多いとき、青は高周波数が多いとき)における複数のニューロンの神経活動パターンの模式図。機械学習法を用いて二つの「感覚経験」情報を最も正しく分類する平面を作製する。(下)この分類でどれほど正確に「感覚経験」情報を復号化できるか計算した。抑制群は対照群と比較して復号精度が低い。「***」有意水準0.1%での有意差あり。
- (D)「感覚経験」情報の保持の回路モデル(後頭頂皮質・視床枕ループ)の模式図。
「感覚経験」情報が後頭頂皮質・視床枕ループで保存されていることが示唆されましたが、前述の通り、この「感覚経験」情報はいつも役に立つわけではなく、現情報試行では正しい判断の妨げにもなります(図1B、C)。実際の状況が変化して「感覚経験」情報が現在の情報とミスマッチすると、感覚経験によるバイアスによって、現情報試行の正しい遂行ができなくなります。それを防ぐためにも「感覚経験」は状況の変化に合わせて動的に更新される必要があります。
その機序を探るために、視床枕の活動を人工的に抑制すると、「感覚経験」情報が更新されるという結果に注目しました(図1B、C、図2B、C)。この結果は、脳内に視床枕の活動を状況に応じて抑制をするシステムがあれば、「感覚経験」の更新が可能になることを示します。視床枕は視床網様核から主な抑制性入力を受けているため、視床網様核がこの更新システムを担っているのではないかという仮説を立て、検証しました(図3A)。
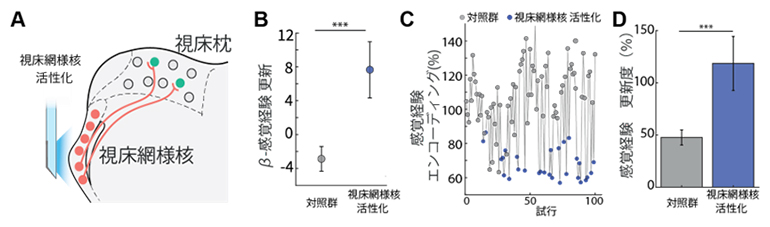
図3 視床網様核の活性化による感覚経験の更新
- (A) 光遺伝学法で視床網様核を活性化すると、視床枕に抑制性入力が提供される(模式図)。
- (B) 行動課題結果から推測される「感覚経験(β‐感覚経験)」情報更新の比較。視床網様核を活性化した群では、活性化しない対照群と比べ、大幅に「感覚経験」情報が更新される。
- (C、D) 後頭頂皮質・視床枕ループに保持された「感覚経験」情報は視床網様核の活性化によって大幅に更新される。「感覚経験」情報がどれくらい神経活動に変換されたか(エンコーディング(%))を解析すると、視床網様核を活性化させた試行で低くなり(C)、「感覚経験」情報の更新度は活性化群が対照群より高い(D)。
まず、視床網様核の活動に「感覚経験」情報の更新能力があるのか調べるために、視床網様核の活動を光遺伝学で活性化させ、「感覚経験」情報が更新されるのかを検証しました。その結果、視床網様核活性化後に視床枕・後頭頂皮質における「感覚経験」情報が更新され、現情報処理能力が上がることが分かりました(図3B、C、D)。このことは、視床網様核が「感覚経験」情報を更新する機能を持つことを示しています。
次に、「感覚経験」情報の動的更新における視床網様核の関与を検証しました。「感覚経験」情報の更新が必要な瞬間を取り出し、その際に視床網様核の活動が上昇している(視床枕の活動が抑制されている)のかを調べました。研究チームが用いた行動課題(図1A)では「感覚経験」情報の更新は、「感覚経験」情報と現情報がミスマッチしている際(最近の5試行で多かった音と現試行で多く流れた音が異なるとき)に起こります(図4A、B)。特に、「感覚経験」情報が強過ぎ、現情報処理ができなくなっていると考えられる[ミスマッチ、不正解試行]において、「感覚経験」情報の更新が必要です。これらの試行における視床枕への抑制性入力を光学的方法(スーパークロメレオン光度測定法[5])で測定し(図4C)、更新と抑制性入力との関連性を調べました。その結果、[ミスマッチ、不正解試行]では、[ミスマッチ、正解試行]に比べて、抑制入力が強くなっていました(図4D左)。一方、「感覚経験」と現情報がマッチしている場合の不正解では、抑制性入力は強くなりませんでした(図4D右)。マッチしている際の不正解は、「感覚経験」と現情報のミスマッチに基づくのではなく、マウスが行動課題に集中していないためだと考えられます。マッチしている場合に正解と不正解での差がないということは、ミスマッチで見られた違いは単に「不正解」をしたからではなく、不正解に導くほどに強く保存された「感覚経験」情報と現情報のミスマッチを検知して更新する過程だと考えられます。
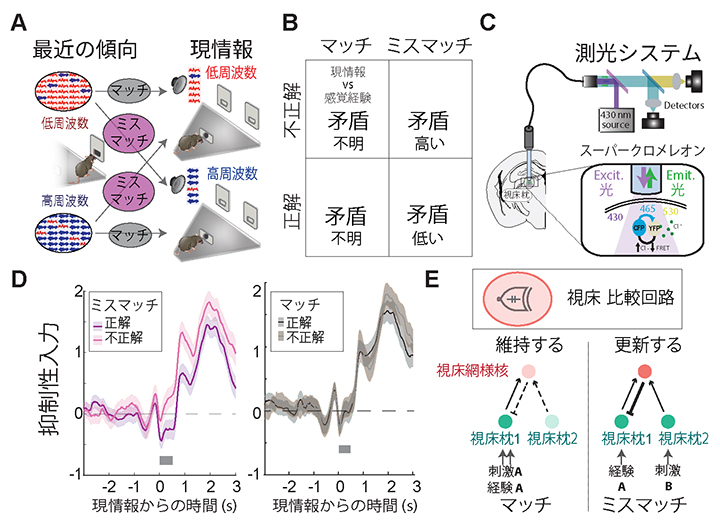
図4 感覚経験が更新されるときに視床網様核の活動は上がる
- (A)行動課題(図1A)中に現れる最近の傾向と現情報のマッチ、ミスマッチ。
- (B)ミスマッチでマウスが不正解した際には「感覚経験」情報と現情報の矛盾が高いことが予測される。
- (C)スーパークロメレオンを用いた測光システムの概要図。スーパークロメレオンに430nmのExcit.光を照射した際のCFPとYFPからのEmit.光の光度変化を比べることによって抑制性入力の強度を測る。
- (D)スーパークロメレオン光度測定法を用いた抑制性入力の測定結果。ミスマッチ不正解で視床枕における抑制性入力が上昇することを示す。
- (E)「感覚経験」情報更新の模式図。結合パターン解析によって、異なる情報を持つ二つの視床枕ニューロンからの視床網様核への入力を発見し、両方の情報が同時に入ってきたときに活性化することが分かった。
そこで、視床網様核が「感覚経験」情報と現情報のミスマッチをどのように検知するのかを検証しました。視床枕から視床網様核へ入力される情報が、「感覚経験」情報と現情報を伝えているのではないかと仮説を立て、各視床網様核ニューロンに入力している視床枕ニューロンの特徴を探りました。
その結果、視床網様核は二つの異なる感覚情報に反応する視床枕ニューロンからの入力を統合し、一つの視床枕ニューロンにフィードバック抑制[6]を行う特徴的な結合パターンを持っていることが明らかになりました(図4E)。このことは、視床網様核が現情報と「感覚経験」情報を照合し、その二つの情報が異なるときに反応する比較器として働くことを示します。
視床網様核が現情報と「感覚経験」情報を照合し、その二つの情報が異なるときに反応する比較器として働く構成は、後頭頂皮質・視床枕・視床網様核回路を模倣したシミュレーションでも証明されました。基本的な回路情報だけを模倣した「後頭頂皮質・視床枕・視床網様核回路」モデルに同じ音刺激判別課題を学習させると、脳と同様の結合パターン(図4E)を持つ視床網様核モデル「ニューロン」が学習後に増えました。この結合パターンを持つ視床網様核モデル「ニューロン」の活動を活性化させると、マウス実験と同様に、人工知能(AI)モデルの経験試行の正答率が下がりました。これらの結果は、「感覚経験」の更新のために生物の進化の中で形成されてきた特定の回路モチーフがAIでも同様につくり上げられ、同じ役割を果たしていることを示します。
今後の期待
「感覚経験」情報に代表される脳の内的情報は、直感や推測の基盤となりますが、状況に応じて更新される一過的な短期記憶である必要があり、ごく一部のみがさまざまなフィルターを経て最終的には長期記憶として定着します。短期記憶から長期記憶への記憶固定化プロセスがうまくいかないと、認知症などで起きたことをすぐ忘れると考えられますがその機序はまだ明らかになっていません。
また、内的情報の更新とは「外の世界の情報」と「脳内のイメージ」処理の切り替えを意味し、この切り替えがうまくいかないと幻聴や思い込みが起きてしまいます。
本研究で「感覚経験」情報が後頭頂皮質・視床枕ループで保存されていることが判明したことにより、この中の一部がどのように取捨選択されて長期記憶になるのかを今後追うことが可能となり、長期記憶への固定化を促進する方法の開発へとつながり、認知症などの治療へ貢献することが期待されます。
また、脳の内的情報が、状況の変化とともにいかに更新されるのか、その計算原理とそれを支える神経回路や、切り替えの神経機序が分かったことで、今後、自分の脳の状態をリアルタイムで確認しながら、この切り替えをまるで瞑想(めいそう)のように自分で制御する「次世代メンタルコントロール」が開発される可能性もあります。
研究チームは、本研究成果が、統合失調症や認知症の治療にとどまらず、個人が持つ能力を引き出し、社会全体のウェルビーイング(幸福)を支える基盤として役立てるように、さらに研究開発に力を注いでいきます。
補足説明
- 1.後頭頂皮質
感覚情報を統合し、空間認識、認知や行動に影響を与える司令塔の役割を持つ大脳皮質の一部。最近の感覚経験情報を保存することが知られている。 - 2.視床枕(ししょうちん)
高次視床核の一部。後頭頂皮質から主要な入力を受けている。主に興奮性ニューロンから成る。他の高次視床核には大脳皮質内情報を安定化する機能があることが知られており、後頭頂皮質内情報の安定化への関与が期待される。 - 3.視床網様核
視床を囲んだ薄い層。他の視床核へ抑制性入力を送り、その活動を抑制する。睡眠や感覚ゲート制御など多くの機能を持つことや統合失調症などにおける機能障害が知られている。 - 4.光遺伝学法
光で活性化されるイオンチャネルやポンプを神経細胞(ニューロン)に発現させ、適切なレーザー光を照射することで、神経活動をミリ秒単位で活性化したり、不活性化したりする技術。 - 5.スーパークロメレオン光度測定法
ニューロン内の塩化物イオン濃度変化を検出するセンサー(スーパークロメレオン)を用いて、抑制性入力を可視化する方法。抑制性神経伝達物質GABAが細胞に取り囲まれる際に上昇する塩化物イオン濃度を測定することで抑制性入力を測ることができる。 - 6.フィードバック抑制
抑制性ニューロンが、自身を活性化するニューロンの活動を抑制し返す(フィードバック)現象。この種の抑制は脳における複雑な情報処理に重要であると考えられている。
研究チーム
理化学研究所 脳神経科学研究センター 認知分散処理研究チーム
チームディレクター シュミット・ルーカス・イアン(Schmitt Lukas Ian)
特別研究員(研究当時)フォスフォード・パトリック(Hosford Patrick)
特別研究員 メイ・ハオ(Mei Hao)
テクニカルスタッフⅠ 田篭 穂沙奈(タゴモリ・ホサナ)
テクニカルスタッフⅠ へイド・キリアン・パトリック(Hayde Cillian Patrick)
特別研究員 アブデルアール・マーナル・サイード(Abdelaal Manal Saeed)
テクニカルスタッフⅠ(研究当時)田篭 帆奈(タゴモリ・ハンナ)
副チームディレクター 中島 美保(ナカジマ・ミホ)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費助成事業基盤研究(B)「知覚経験表象の戦略的更新を可能にする視床皮質ネットワークの解明(研究代表者:シュミット・ルーカス、23K24000)」による助成を受けて行われました。
原論文情報
- Patrick Steven Hosford, Hao Mei, Hosana Tagomori, Cillian Patrick Hayde, Manal Saeed Abdelaal, Hanna Tagomori, Miho Nakajima, Lukas Ian Schmitt, "Control of Representation Updating by Higher-Order Thalamus Enables History-Based Decision-Making", Neuron, 10.1016/j.neuron.2026.04.045
発表者
理化学研究所
脳神経科学研究センター 認知分散処理研究チーム
チームディレクター シュミット・ルーカス・イアン(Schmitt Lukas Ian)
特別研究員(研究当時)フォスフォード・パトリック(Hosford Patrick)
特別研究員 メイ・ハオ(Mei Hao)
副チームディレクター 中島 美保(ナカジマ・ミホ)
 フォスフォード・パトリック
フォスフォード・パトリック
 メイ・ハオ
メイ・ハオ
 シュミット・ルーカス・イアン
シュミット・ルーカス・イアン
発表者のコメント
視床の本質を少しでも解き明かす一助となれたこと、そしてこの素晴らしい多角的な視点を持ったチームの一員になれたことを光栄に思います。私たちの問いを解決するには、材料工学、電気工学、計算解析、光学、そして細胞生物学の専門知識が必要不可欠でした。この困難な課題に一緒に立ち向かい、多大なる貢献をしてくれた共著者の方々に深く感謝いたします。(フォスフォード・パトリック)
過去の経験は、私たちがどのような人間であるかを形作り、現在の行動に影響を与えます。こうした経験には、最近取り入れたばかりの感覚情報も含まれます。感覚が曖昧な信号を捉えたときなど、困難な状況に対処するために、私たちの脳はこうした直近の情報をどのように活用しているのでしょうか。この問いに対して、たとえわずかであっても答えを出そうと、才能あふれる共著者の方々と共に研究に取り組めたことを大変光栄に思います。(メイ・ハオ)
私はかねてより、世界の事象からパターンを感知し、推論を導き出す「直感」という概念に深く魅了されてきました。それは探偵が証拠をつなぎ合わせるプロセスにも似ていますが、人間の脳は形式的な論理を介さずとも、多様なパターンを絶えず抽出し、その変遷を把握する優れた能力を有しています。この直感は洞察の源泉として強力な武器となる一方、現代の情報過多な環境下では、その機能が阻害されかねないという危うさもはらんでいます。故に、本研究が試みた脳の推論プロセスの科学的解明は極めて重要であり、将来的にはテクノロジーを通じてこの能力を研さんする、新たなパラダイムの創生に寄与するものと確信しています。(シュミット・ルーカス・イアン)
報道担当
理化学研究所 広報部 報道担当
お問い合わせフォーム