理化学研究所(理研)生命医科学研究センター 免疫転写制御研究チームの谷内 一郎 チームディレクター、東京医科大学 分子病理学分野の黒田 雅彦 主任教授らの国際共同研究グループは、外来性抗原に対する免疫寛容[1]が樹立されるのに必須の末梢誘導性制御性T細胞[2]を産み出す抗原提示細胞[3]の分化制御機構を明らかにし、この細胞を人為的に操作することで末梢誘導性制御性T細胞を増やせることを明らかにしました。
本研究成果は、外来性抗原への過剰な免疫応答が原因で発症する炎症性疾患や食物アレルギーに対する治療法、移植医療分野での拒絶反応抑制法の開発につながる成果といえます。
今回、国際共同研究グループは転写因子であるRunx1タンパク質[4]の発現操作により、Thetis細胞[5]と名付けられた特異な抗原提示細胞の分化が増強され、その結果、末梢誘導性制御性T細胞を増やすことが可能であることを発見しました。
本研究は、科学雑誌『Nature Immunology』オンライン版(6月19日付:日本時間6月19日)に掲載されました。
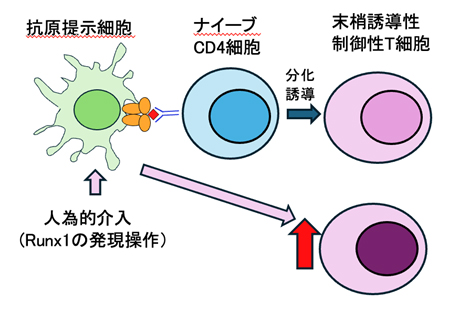
抗原提示細胞に介入し制御性T細胞を増やす(赤矢印は増えることを意味)方法を開発
背景
制御性T細胞は免疫反応を抑制する能力を有する特別なCD4陽性T細胞です。その発見の功績に対し坂口 志文 博士に2025年のノーベル生理学・医学賞が贈られたことは記憶に新しく、制御性T細胞を活用した新たな治療法の開発に期待が寄せられています。
制御性T細胞には、胸腺というT細胞がつくられる臓器で発生するものと、末梢組織でナイーブCD4細胞[6]から分化するものの2種類があることが知られています。
胸腺由来の制御性T細胞は、自己抗原を認識するT細胞抗原受容体を発現する細胞の一部が、Foxp3という制御性T細胞のマスター転写因子[7]を発現することで発生します。そのため、胸腺由来の制御性T細胞ができないと、自己抗原に対する免疫応答が抑制されず激しい自己免疫病が発症します。
一方で、ヒトの身体の中では、腸管を代表として常在性細菌叢(さいきんそう)が共生し、腸管には食物由来の成分も多数存在します。このような常在性細菌や食物由来の抗原は自己抗原ではなく外来性の抗原です。ヒトの身体にはこのような外来性抗原に対して過剰な免疫応答が起こらない仕組みが備わっていて、これは外来性抗原に対する免疫寛容と呼ばれます。
最近の研究から、この免疫寛容には、末梢組織でナイーブCD4細胞から分化する末梢誘導性制御性T細胞が必須であることが分かってきました。この末梢誘導性制御性T細胞は、Foxp3転写因子に加えRorγt転写因子も発現することからRorγt陽性制御性T細胞とも呼ばれます。末梢誘導性制御性T細胞の産生には、外来性抗原の情報をナイーブCD4細胞に提示して末梢誘導性制御性T細胞への分化を誘導する役割を持つ、いわば末梢誘導性制御性T細胞の親のような細胞が存在するはずです。末梢誘導性制御性T細胞の「母なる細胞」が見つかり、その数や機能を操作することができれば、末梢誘導性制御性T細胞を増やすことが可能になると考えられることから、その発見は極めて激しい競争状態となっていました。その発見のためには、この母なる細胞の欠損により末梢誘導性制御性T細胞が産生されないマウスが有用な研究材料となります。
研究手法と成果
理研免疫転写制御研究チームでは、遺伝子変異マウスを用いて免疫細胞分化におけるRunx/Cbfβ転写因子複合体(Runxタンパク質とCbfβタンパク質がヘテロ(異種)2量体を形成したもの)の機能を長年研究してきました。Cbfβ遺伝子からはRNAスプライシング[8]によりCbfβ1とCbfβ2というC末端[9]のアミノ酸構造が異なるCbfβタンパク質が産生されます。
Cbfβ1とCbfβ2がそれぞれ特異な機能を有するか調べるために、国際共同研究グループはCbfβ遺伝子に変異を導入してCbfβ2を特異的に欠損するマウス(Cbfβ2欠損マウス)を作製しました。その結果、Cbfβ2欠損マウスは3カ月齢という若い年齢で大腸炎を自然発症することが分かりました(図1A)。
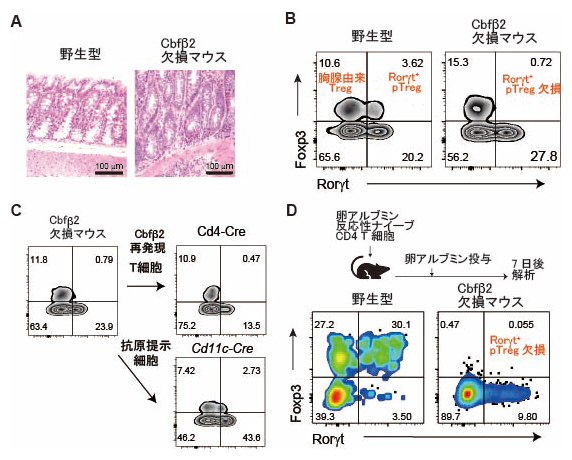
図1 Cbfβ2欠損マウスは抗原提示細胞の問題で末梢誘導性制御性T細胞が欠損
- (A)野生型マウスとCbfβ2欠損マウスの大腸の組織像。Cbfβ2欠損マウスでは大腸炎が起こっている。
- (B)腸のT細胞のフローサイトメトリー(1個1個の細胞にレーザー光を照射し、その散乱光や蛍光を測定することで、細胞の情報を取得したり、特定の細胞を分取したりする技術)解析例。Cbfβ2欠損マウスでは末梢誘導性制御性T細胞(Rorγt+ pTreg)の細胞集団が欠失している。
- (C)Cbfβ2の再発現実験の結果。抗原提示細胞でCreタンパク質を発現するCd11c-Creマウスでは末梢誘導性制御性T細胞(Rorγt+ pTreg)の細胞集団が回復する。一方、T細胞でのトランスジェニックCbfβ2タンパク質を発現するCd4-Creマウスでは、末梢誘導性制御性T細胞(Rorγt+ pTreg)の細胞集団は回復しなかった。
- (D)ナイーブCD4 T細胞を移入する実験結果。Cbfβ2欠損マウスでは移入したナイーブCD4 T細胞は末梢誘導性制御性T細胞(Rorγt+ pTreg)に分化できない。
一方、末梢誘導性制御性T細胞は大腸炎の発症抑制に重要な働きをすることが別の研究注1)から判明していたので、国際共同研究グループはCbfβ2欠損マウスに末梢誘導性制御性T細胞が存在するかを調べました。その結果、腸管の固有粘膜層に胸腺由来のRorγt陰性制御性T細胞(Foxp3+Rorγt- pTreg)は存在しているが、末梢誘導性のRorγt陽性制御性T細胞(Foxp3+Rorγt+ pTreg)はほとんど存在しないことが分かりました(図1B)。
Runx/Cbfβ転写因子複合体はT細胞でも発現しFoxp3遺伝子の発現調節に関与します。そこで国際共同研究グループは、Cbfβ2欠損マウスでの末梢誘導性制御性T細胞(Rorγt+ pTreg)の欠失がT細胞の内因的な原因によるのか、T細胞以外に原因があるのか調べることにしました。
その方法として、遺伝学的手法を用いT細胞やそれ以外の免疫細胞でCbfβ2タンパク質の発現を回復することで、末梢誘導性制御性T細胞が分化するかどうかを調べる実験を行いました。実際には、Creタンパク質[10]の発現によりSTOP配列[11]を除去して、トランスジェニックCbfβ2タンパク質の発現が誘導される実験系を構築し、Creタンパク質をどの細胞で発現させるか調整することで、特定の細胞にトランスジェニックCbfβ2タンパク質を発現させることができます。その結果、T細胞でのトランスジェニックCbfβ2タンパク質の発現では、末梢誘導性制御性T細胞の分化は回復しませんでした(図1C)。この結果は、末梢誘導性制御性T細胞の消失の原因がT細胞でのCbfβ2欠損ではないことを示します。
さらに国際共同研究グループは、卵のアルブミンタンパク質に反応するナイーブCD4 T細胞を野生型マウスとCbfβ2欠損マウスに移入し、卵のアルブミンを経口投与することで末梢誘導性制御性T細胞に分化するか調べました。その結果、野生型マウスでは移入したナイーブCD4 T細胞から末梢誘導性制御性T細胞が発生したのに対し、Cbfβ2欠損マウスでは末梢誘導性制御性T細胞は発生しませんでした(図1D)。この結果は、Cbfβ2欠損マウスでは、ナイーブCD4T細胞に外来性抗原の情報を伝えて末梢誘導性制御性T細胞の分化を誘導する抗原提示細胞に問題があることを示します。Cd11c-Creマウスは、抗原提示細胞を含む細胞集団でCreタンパク質を発現しますが、このCd11c-Creを用いてトランスジェニックCbfβ2タンパク質をCbfβ2欠損マウスに発現させると末梢誘導性制御性T細胞分化が回復しました。
こうしたことから、国際共同研究グループは次に、Cbfβ2欠損マウスでは、末梢誘導性制御性T細胞の分化を誘導する抗原提示細胞にどのような問題があるか調べることにしました。
Thetis細胞は末梢誘導性制御性T細胞の分化を誘導する抗原提示細胞として2022年に発見された細胞で、少なくとも4種類のサブセット(TC-I、TC-II、TC-III、TC-IV)があり、最近の研究注2)からはTC-IVが末梢誘導性制御性T細胞の分化誘導に重要であることが分かってきています。そこで国際共同研究グループは、フローサイトメトリーという細胞解析法でThetis細胞を同定する方法をブラウン・クリソセミス博士から学び、Cbfβ2欠損マウスを調べました。その結果、Cbfβ2欠損マウスではTC-II、TC-III、TC-IVが欠失していることが分かりました(図2中)。Cbfβ2欠損マウスではCd11c-Creを用いたトランスジェニックCbfβ2タンパク質発現により末梢誘導性制御性T細胞の分化が回復します。しかし、このマウスでThetis細胞分化はどのようになっているか解析したところ、TC-IVサブセットの分化のみが回復していました(図2下)。この結果はTC-IVサブセットの回復のみで末梢誘導性制御性T細胞の分化誘導には十分であることを示す、初めての遺伝学的証拠です。
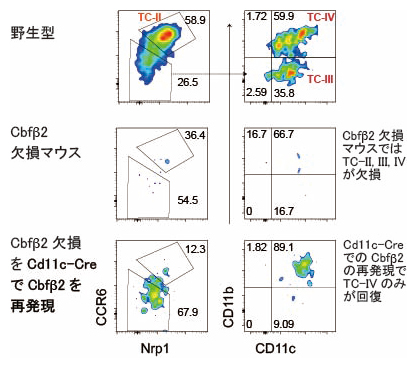
図2 Cbfβ2欠損マウスでのTCsの欠損とCbfβ2再発現でのTC-IVの回復
フローサイトメトリーによるThetis細胞(TCs)の解析例。Cbfβ2欠損マウスではTC-II、TC-III、TC-IVが欠損しており、Cd11c-Creマウスを用いたCbfβ2の再発現によりTC-IVのみが回復した。
CbfβはRunxタンパク質と2量体を形成し機能します。哺乳動物では3種類のRunx遺伝子からそれぞれRunx1、Runx2、Runx3の3種類のRunxタンパク質が産生されます。そこで国際共同研究グループはどのRunxタンパク質がThetis細胞の発生に関与するか調べることにしました。
Runx3遺伝子の変異によりマウスが大腸炎を発症することが報告されていたこともあり、国際共同研究グループはまず、Runx3に着目し、新しいRunx3変異マウスであるRunx3-WRPWマウス注3)を調べました。Runx3-WRPWマウスでは、Runx3タンパク質のC末端にあるWRPYモチーフ(構造)内のチロシン(Y)残基がトリプトファン(W)残基に置換されていることで、本来のRunx3の機能が阻害されます。Cbfβ2欠損マウスと同様に、Runx3-WRPWマウスは大腸炎を自然発症し、末梢誘導性制御性T細胞が欠失し、さらにTC-II、TC-III、TC-IVも欠失が観察されました。
続いて国際共同研究グループはCd11c-Creトランスジェニックマウスを用いた条件付きRunx3遺伝子破壊の影響を調べました。その結果、この実験系ではRunx3遺伝子の単独破壊で末梢誘導性制御性T細胞の欠失は見られず、Runx3遺伝子に加えRunx1遺伝子を同時に破壊することで、末梢誘導性制御性T細胞とTC-III、TC-IVの欠失が同時に起こることを発見しました(図3左の中)。
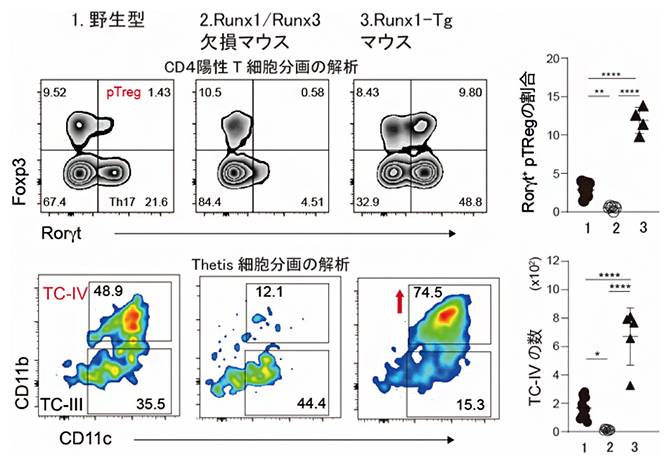
図3 トランスジェニックRunx1によるTC-IV、末梢誘導性制御性T細胞の増加
Runx1とRunx3をCD11c-Creで不活性化すると、腸のCD4+T細胞集団の中で末梢誘導性制御性T細胞(Rorγt+ pTreg)が消失し、Thetis細胞ではTC-IVが欠損する。トランスジェニックRunx1の発現により、TC-IVや末梢誘導性制御性T細胞は回復するばかりでなく、増加した。右の図の1、2、3はそれぞれ、1.野生型、2.Runx1/Runx3欠損マウス、3.Runx1-Tgマウスを示す。「*」有意水準5%での有意差あり、「**」有意水準1%での有意差あり、「****」有意水準0.01%での有意差あり。
国際共同研究グループは、Creタンパク質の発現によりRunx1タンパク質の発現を誘導できるマウスを保有していたので、Cd11c-Creトランスジェニックマウスにより内在性のRunx1遺伝子とRunx3遺伝子を破壊すると同時に、トランスジェニック(Tg)Runx1タンパク質の発現を誘導するマウス(Runx1-Tg)を作製しました。予想通りトランスジェニックRunx1タンパク質の発現によりTC-III、TC-IVの欠失は回復しました(図3左の右)。詳細に観察すると、Runx1-TgでTC-III、TC-IVの細胞数は増加し、特にTC-IVの増加が顕著でした(図3右の下)。さらに末梢誘導性制御性T細胞の分化を見てみると、トランスジェニックRunx1タンパク質の発現により、腸管内の末梢誘導性制御性T細胞の割合は正常の野生型マウスと比べ3倍程度増加していることが分かりました(図3右の上)。
この結果は、Cd11c-CreによるトランスジェニックRunx1タンパク質の発現操作という人為的介入により、TC-IVという抗原提示細胞の分化・機能を増強することが可能であり、その結果としてマウス生体内で末梢誘導性制御性T細胞を増やすことができることを示しています。
- 注1)Caspar Ohnmacht et al. The microbiota regulates type 2 immunity through RORγt+ T cells, Science, 9 Jul 2015.
- 注2)Vanja cabric et al. A wave of Thetis cells imparts tolerance to food antigens early in life. Science, 15 May 2025.
- 注3)2026年2月19日プレスリリース「免疫細胞の運命を決める"リン酸化スイッチ"を発見」
今後の期待
本研究は、Runx/Cbfβ転写因子がThetis細胞の発生に必須であることを明らかにし、Thetis細胞の中でもTC-IVサブセットが、末梢誘導性制御性T細胞の分化に必須の抗原提示細胞であるという遺伝学的な証拠を示したものです。またRunx転写因子の発現を操作することで、TC-IVサブセットの分化が増強され、その結果腸管内の末梢誘導性制御性T細胞の数を増加させることが可能であることを明らかにしました。抗原提示細胞を操作することで、マウス生体内で末梢誘導性制御性T細胞を増やす方法は世界で初めてのものであり、今後は炎症性疾患や食物アレルギーに対する新規治療法開発への応用が期待されます。さらに外来性抗原への免疫寛容の成立という視点では、移植医療において異種臓器への免疫寛容の誘導への応用も期待されます。
補足説明
- 1.免疫寛容
特定の抗原に対する免疫応答が抑制または欠如している免疫の仕組みやその状態。 - 2.末梢誘導性制御性T細胞
制御性T細胞は免疫の働きが行き過ぎないように整えて、体のバランスを保つ"調整役"の細胞。Foxp3転写因子を発現するCD4+T細胞として同定され、英語での略称でTreg(Tレグ)と呼ばれることもある。末梢誘導性制御性T細胞は、末梢組織でナイーブCD4細胞([6]参照)から分化し、FoxP転写因子に加えRorγt転写因子を共発現する特徴を有する。 - 3.抗原提示細胞
抗原を取り込んでMHCと呼ばれる細胞表面分子に載せ、T細胞に抗原の特徴を提示する細胞。 - 4.Runx1タンパク質
Runxタンパク質は進化上、線虫から保存されている転写因子の一種で、細胞の分化や増殖などの重要な生物学的プロセスを制御している。哺乳動物ではRunx1、Runx2、Runx3の3種類があり、Runx1は造血系の形成に必須であることが知られている。 - 5.Thetis細胞
Rorγt転写因子の発現を特徴とする抗原提示細胞。末梢誘導性制御性T細胞(Rorγt+pTreg)分化を誘導する抗原提示細胞は、2020年頃から世界で多くの研究室が探している細胞で、その探索は非常に厳しい競争の状況であったが、2022年にブラウン・クリソセミス博士が、その候補として新しいタイプの抗原提示細胞を見つけ、ギリシャ神話の海の女神テティス(Thetis)から名前を取りThetis細胞(Thetis Cellsを略してTCsと表記することもある)と名付けた。 - 6.ナイーブCD4細胞
抗原による刺激を受けておらず、活性化していないCD4陽性T細胞。抗原提示細胞から抗原刺激を受けると活性化されエフェクターCD4細胞に分化する。 - 7.マスター転写因子
特定の細胞の「運命」や性質を決定付ける中心的な役割を持ち、多数の標的遺伝子群の発現を統括的に制御する転写因子。 - 8.RNAスプライシング
DNA遺伝情報がmRNAに転写され生成されたRNA前駆体の一部が、切断されて除かれた後、残りの部分が再結合する反応。 - 9.C末端
タンパク質はアミノ酸同士が脱水縮合して形成されたポリマーであり、隣接するアミノ酸は、それぞれのアミノ基とカルボキシ基がペプチド結合をしている。このポリマーの末端のフリーのアミノ基側をN末端、カルボキシ基側をC末端と呼ぶ。 - 10.Creタンパク質
loxPと呼ばれる特異なDNA配列を認識し、loxP配列同士間での部位特異的DNA組換えを起こす酵素活性を持つタンパク質。 - 11.STOP配列
遺伝子が読み取られるのを途中で強制的にストップさせる、特殊なブレーキ役となるようなDNA配列。遺伝子の転写をストップさせるpolyA(pA)配列がよく使われる。この配列の両端に「loxP」を配置させると、Cre酵素によってSTOP配列が切り取られ、その先の遺伝子が転写されるようになり、Cre発現細胞で遺伝子の発現スイッチをオンにできる。
国際共同研究グループ
理化学研究所 生命医科学研究センター
免疫転写制御研究チーム
チームディレクター 谷内 一郎(タニウチ・イチロウ)
研究員 小川 ちひろ(オガワ・チヒロ)
特別研究員(研究当時)ゾウ・チェンチェン(Zou Chengcheng)
基礎科学特別研究(研究当時、現 客員研究員)山下 基(ヤマシタ・モトイ)
大学院リサーチ・アソシエイト(研究当時、現 特別研究員)原田 淳司(ハラダ・ジュンジ)
空間免疫制御理研ECL研究ユニット
理研ECL研究ユニットリーダー 佐藤 尚子(サトウ・ナオコ)
東京医科大学 分子病理学分野
主任教授 黒田 雅彦(クロダ・マサヒコ)
スローンケタリングがんセンター(米国)
准教授 ブラウン・クリソセミス(Brown Chrysothemis)
研究員 パーク・タイラー(Park Tyler)
博士課程大学院生 パウカイザ・ヨスリン(Paucar Iza Yoselin)
研究支援
本研究は、日本学術振興会(JSPS)科学研究費助成事業新学術領域研究(研究領域提案型)「血球系細胞分化過程での非ゲノム情報複製機構の解明(研究代表者:谷内一郎)」、同基盤研究(B)「C末端構造を介したRunx転写因子の機構制御機構の解明(研究代表者:谷内一郎)」、同基盤研究(A)「末梢性免疫寛容樹立機構の解明と介入法の開発(研究代表者:谷内一郎)」、同基盤研究(C)「Runx依存的Thetis細胞分化制御機構の解明(研究代表者:小川ちひろ)」による助成を受けて行われました。
原論文情報
- Chihiro Ogawa, Chengcheng Zou, Yoselin A Paucar Iza, Motoi Yamashita, Tyler Park, Junji Harada, Naoko Satoh-Takayama, Masahiko Kuroda, Chrysothemis C Brown and Ichiro Taniuchi, "Runx/Cbfβ regulates the development of tolerogenic Thetis cells", Nature Immunology, 10.1038/s41590-026-02566-8
発表者
理化学研究所
生命医科学研究センター 免疫転写制御研究チーム
チームディレクター 谷内 一郎(タニウチ・イチロウ)
東京医科大学 分子病理学分野
主任教授 黒田 雅彦(クロダ・マサヒコ)
 谷内 一郎
谷内 一郎
発表者のコメント
Cbfβ2欠損マウスは、私が20年以上前に、独立研究者になり、初めて作製したマウスです。大腸炎を自然発症することはすぐに気づきましたが、その原因は長く解明できませんでした。他の研究者が末梢誘導性制御性T細胞を発見し、2022年にThetis細胞が発見されたことでようやくRunx/Cbfβ変異マウスが大腸炎を起こす原因を突き止めることができました。トランスジェニックRunx1の発現でTC-IVや末梢誘導性制御性Tが回復するばかりでなく、増加することは予期せぬ発見でしたが、この発見は治療応用への大きな可能性を示すもので、長年粘って研究してきたことへご褒美を頂いた気分です。(谷内 一郎)
報道担当
理化学研究所 広報部 報道担当
お問い合わせフォーム
東京医科大学 企画部 広報・社会連携推進室
Tel: 03-3351-6141(代表)
Email: d-koho@tokyo-med.ac.jp